

Article | 05/06/2024
Les enveloppes terrestres externes et leurs relations avec la biosphère au Protérozoïque
05/06/2024
Résumé
Stase biogéochimique et Évènement d'Oxygénation Néoprotérozoïque (NOE) – Glaciations extrêmes et évolution des eucaryotes.
Table des matières
Le Protérozoïque correspond à l'éon (plus haute division stratigraphique) situé entre l'Archéen et le Phanérozoïque, de −2,5 Ga à −542 Ma. Malgré une durée de deux milliards d'années, les données bibliographiques concernant cette période sont rares. Le Protérozoïque est souvent présenté comme une période de transition entre l'Archéen – à la tectonique mixte verticale (sagduction) et horizontale (avec des plaques moins rigides que les plaques actuelles), aux enveloppes externes anoxiques, avec peu de traces de vie fossilisables (stromatolithes, BIFs – banded iron formations ou fers rubanés) – et le Phanérozoïque – avec les enveloppes externes oxygénées et sa vie « apparente ». Pendant le Protérozoïque, les enveloppes externes sont modérément oxygénées, à l'exception de l'océan profond, anoxique la plupart du temps (Kump, 2008 [15]), alors que la tectonique des plaques correspond à la tectonique actuelle, de même que les roches de la série calco-alcaline (Martin et al., 2010 [20]). D'un point de vue évolutif, le Protérozoïque peut être considéré comme l'éon au début duquel les eucaryotes apparaissent et se développent discrètement (les stromatolithes restent fréquents, les fossiles d'eucaryotes unicellulaires sont rares, souvent des « Acritarches », probablement affiliés aux Dinoflagellés) et à la fin duquel les métazoaires se manifestent dans les faunes de Doushantuo (Xiao et al., 2000 [31]) et Ediacara4 (Xiao et Laflamme, 2009 [32] ; Langlois, 2018 [17]) (Dickinsonia, Charnodiscus, Tribrachidium…). Des formes de vie inconnues ont pu apparaitre et disparaitre sans laisser de traces dans le registre fossile, si ce n'est qu'à l'état de molécules qui commencent à être détectées grâce à des innovations techniques (extraction à partir de matériel complexe, chromatographies très sensibles – Brocks et al., 2023 [4]). Le Protérozoïque est encadré par deux épisodes d'oxygénation, le premier juste après la fin de l'Archéen (Grand Évènement Oxydant, −2,45 à −2,2 Ga) accompagné du premier épisode de snowball Earth (Terre boule de neige) vers 2,4 Ga, et le suivant à son extrémité terminale, de −720 Ma à −580 Ma, précédé par des glaciations extrêmes (snowball Earths sturtienne et marinoenne, épisode Gaskiers) qui correspondent au Cryogénien. À la fin du Cryogénien, les teneurs en oxygène de l'atmosphère sont comparables à celles reconstruites tout au long du Phanérozoïque, et permettent l'établissement de faunes de type Ediacara. Entre ces deux épisodes, on n'observe pas de grande glaciation, les traces de vie restent discrètes, pas d'évènement tectonique majeur. Cette période, située entre −1,88 Ga (derniers BIFs anciens) et −720 Ma (début du Cryogénien) est surnommée le « boring billion » (milliard ennuyeux).
Nous allons voir comment les enveloppes externes et le monde vivant ont évolué depuis la fin du Grand Événement Oxydant (GOE) jusqu'au début du Phanérozoïque. Les méthodes de reconstitution de l'état rédox des enveloppes externes seront présentées, les causes de la stase biogéochimique protérozoïque (teneur en oxygène autour de 2 % dans l'atmosphère) et de l'événement d'oxygénation néoprotérozoïque (NOE) seront discutées.
Les différents indicateurs de la teneur en oxygène des enveloppes externes
Les affleurements les plus anciens sont rares et potentiellement soumis à une longue histoire d'altération, de métamorphisme ou d'oxydation, étant en contact avec une atmosphère contenant 20 % d'oxygène. Les indices d'oxygénation sont indirects et illustrent la créativité des géochimistes et des géologues qui arrivent à déduire des informations cruciales d'un ensemble épars d'observations. Par exemple, l'absence d'uraninite et de pyrite détritiques après le Grand Évènement Oxydant (GOE) implique une teneur en oxygène au moins égale à 0,2 % (Thomas, 2022 [30]). Les charbons de bois, présents à chaque époque dans des sédiments continentaux depuis le Silurien (420 Ma) attestent d'une teneur en oxygène comprise entre 12 % et 32 %. En dessous de 12 %, les flammes ne peuvent pas persister ; au-dessus de 32 %, les feux de forêt continuels empêcheraient l'établissement de forêts. Les indicateurs paléontologiques de la teneur en oxygène sont malheureusement peu robustes pour de longues durées (signal des ostracodes Podocopides (Horne et al., 2011 [14]), taille des insectes (Clapham et Karr, 2012 [8]). Les méthodes géochimiques vont concerner des rapports isotopiques (soufre, chrome), des teneurs en différents oxydes (fer, manganèse).
# Le fractionnement indépendant de la masse des isotopes du soufre et la présence d'une couche d'ozone
Sur la Terre moderne, les isotopes d'un même élément (par exemple 32S, 33S, 34S, 36S) se répartissent inégalement entre les différentes phases ou substances en présence (on parle de fractionnement isotopique) en fonction de leur masse pendant des processus physiques ou biologiques selon des règles précises. Par exemple, la réduction des sulfates (SO42−) par les bactéries sulfato-réductrices montre une préférence pour le 32S (16 protons + 16 neutrons) plus léger que les isotopes 33S et 34S [fractionnement cinétique comparable à celui observé pour 12C et 13C lors de la photosynthèse], ce qui conduit à la formation d'une pyrite sédimentaire (dans des environnements anoxiques) enrichie en 32S par rapport au gypse CaSO4.2 H2O formé à partir des mêmes eaux. On mesure les δ33S et δ34S par rapport à l'isotope 32.
Les règles de fractionnement dépendant de la masse sont :
δ33S = 0,515 × δ34S ; dans ce cas, en utilisant la notation Δ[1], Δ33S = 0 (i.e., le fractionnement de 33S par rapport à 32S est la moitié du fractionnement de 34S par rapport à 32S comme la différence de masse entre l'isotope 33 et l'isotope 32 est la moitié de celle entre l'isotope 34 et l'isotope 32 du soufre)
et également
δ36S = 1,9 × δ34S ; dans ce cas Δ36S = 0 (i.e., le fractionnement de 36S par rapport à 32S est approximativement le double du fractionnement de 34S par rapport à 32S, pour les mêmes raisons).
Il existe un fractionnement des isotopes indépendant de la masse. Parmi les processus qui conduisent à ce fractionnement, les interactions entre rayonnements très énergétiques et soufre d'origine volcanique sont fondamentaux. Les UV « durs » (à courte longueur d'onde donc très énergétiques) dans l'atmosphère peuvent entrainer la réaction de dismutation du dioxyde de soufre SO2 en sulfates et soufre natif S° :
3 SO2 + 3H2O → 2 HSO4− + S° + 2H+.
Cette réaction fractionne les isotopes de manière indépendante de la masse (Δ33S ≠ 0, Δ36S ≠ 0) pour chacune des phases. En 2000, Farquhar et al. [11] ont mesuré la composition isotopique du soufre dans les sulfates (gypse) et sulfures archéens et modernes. On voit dans la figure extraite de leur publication, de manière non ambigüe, que le fractionnement des isotopes du soufre est indépendant de la masse (donc issu d’un effet photochimique) de 4 Ga jusqu'à 2,5 Ga ; après 2 Ga, le fractionnement des isotopes est dépendant de la masse. Une “zone grise” entre 2,5 et 2 Ga montre un fractionnement presque dépendant de la masse.
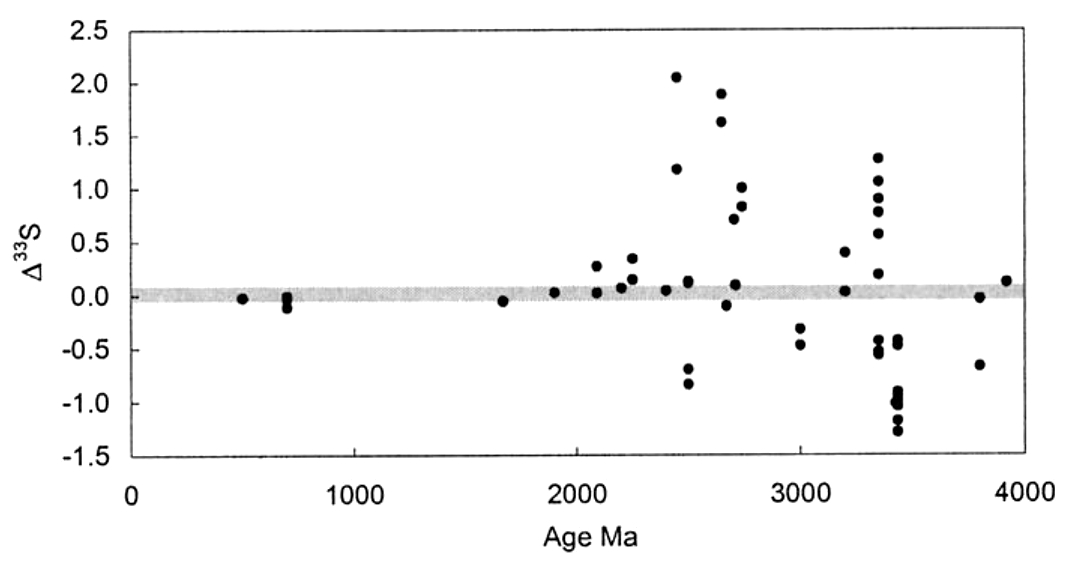
Source - © 2000 D’après Farquhar et al. [11]
Figure. Fractionnement isotopique du soufre depuis 4 Ga
Le fractionnement indépendant de la masse (Δ33S ≠ 0) chute après 2,45 Ga et reste quasi nul après 2,2 Ga. Il y a donc un phénomène qui empêche les UV durs de fractionner les isotopes du soufre, probablement de l'ozone (qui absorbe ces radiations). Si de l'ozone est présent dans l'atmosphère, c'est qu'il y a de l'oxygène.
Le fractionnement indépendant de la masse ne peut avoir lieu que si des UV durs traversent l'atmosphère. Si de l'ozone est présent dans l'atmosphère, les UV durs sont absorbés, il n'y a alors plus de fractionnement indépendant de la masse. Pour que de l'ozone soit présent, il faut du dioxygène dans l'air. Le GOE s'est donc produit entre 2,4 et 2 Ga.
# Isotopes du chrome et oxygénation de l'atmosphère
Le chrome possède quatre isotopes de masses 50, 52, 53 et 54. Le comportement géochimique du chrome est très sensible aux conditions rédox : l'altération en conditions oxydantes produit la forme hexavalente Cr(VI) ; l'oxydation de la forme trivalente Cr(III) sur les continents est accompagnée d'un fractionnement isotopique qui enrichit la phase mobile Cr(VI) en isotope lourd.
Comme l'histoire rédox de la roche peut modifier les rapports Cr(VI)/Cr(III), on mesure les rapports isotopiques, un enrichissement en 53Cr indiquant du chrome oxydé à l'origine (pourvu qu'il n'y ait pas eu de migration). Les données isotopiques (δ53Cr mesurant le fractionnement de 53Cr par rapport à 52Cr) indiquent une accumulation de chrome hexavalent (Cr(VI) oxydé) à la surface des océans entre 2,8 et 2,6 Ga avec une augmentation juste avant le Grand Événement Oxydant10 (Frei et al., 2009 [12]). Dans des BIFs de 1,88 Ga, les isotopes du chrome ne sont pas fractionnés, ce qui indique un déclin de la teneur en oxygène atmosphérique. Enfin, à la fin du Néoprotérozoïque, on observe de très forts fractionnements qui prouvent une augmentation de l'oxygénation de surface à cette époque, qui a sans doute stimulé le développement de la vie macroscopique multicellulaire.
# Présence de fer dans les paléosols
Dans un environnement réducteur, le Fe2+ n'est pas oxydé, il est alors mobile (soluble) lors de l'altération/érosion de la roche mère. Dans un environnement oxydant, le Fe2+ s'oxyde en Fe3+ et reste sur place, le plus souvent sous forme d'oxy-hydroxydes de fer (FeOOH, Fe(OH)3 et tous les dérivés). Des travaux variés ont été entrepris sur les paléosols archéens, notamment en Afrique du Sud, au Botswana, au Mozambique. Les sols antérieurs à 2,44 Ga ont perdu beaucoup de fer (par rapport à la roche mère) ; les sols dont l'âge est compris entre 2,4 et 2,2 Ga présentent des résultats ambigus ; des Red Beds (sols dans lesquels la perte de fer est minimale) de 2,2 Ga surmontent le tout (Rye et Holland, 1998 [23]).
# Précipitation bactérienne du manganèse
Le manganèse est un métal utilisé dans la fabrication de certains aciers et dans celle des batteries au lithium. Ce métal présente trois états d'oxydation, Mn(II), Mn(III) et Mn(IV), les deux derniers précipitant facilement sous forme d'oxydes. La présence d'O2 est nécessaire pour oxyder Mn(II), cette oxydation se faisant grâce à l'action de bactéries, du fait d'un blocage cinétique. Au sein des sédiments anoxiques, les oxydes Mn(III,IV) peuvent être réduits et remobilisés par l'oxydation de matière organique médiée par des bactéries, ou par des réactions abiotiques avec des sulfures ou du Fe(II). Les principales ressources en manganèse se sont formées lors de l'oxygénation de l'atmosphère, les plus anciennes datant de 2,4 Ga (mines du Kalahari en Namibie). À l'Ouest de l'Australie, au sein du craton de Pilbara, un bassin sédimentaire tardi-orogénique, le bassin d'Edmund, contient des ressources en manganèse considérables. Ces dépôts sont stratiformes, et interprétés comme liés à l'oxydation de sédiments plus anciens (Spinks et al., 2023 [26]). Leur datation à 1,4-1,1 Ga montre une augmentation temporaire de la teneur en oxygène atmosphérique.
Cette liste n'est évidemment pas exhaustive, les comportements géochimiques du molybdène, qui “adore” les sulfures, ou du vanadium, sont également informatifs. Malgré la diversité des indicateurs de l'oxygénation des enveloppes externes, il n'existe pas d'enregistrement de cette oxygénation continu et dénué d'incertitude. Les résultats présentés ci-dessous sont donc un consensus provisoire dans la communauté scientifique.
L'évolution chimique de l'océan au cours du Protérozoïque
Les affleurements datant du Protérozoïque sont un peu plus abondants que les affleurements de l'Archéen. Par exemple, en France métropolitaine, dans le bloc de Saint-Brieuc (Massif Armoricain), affleurent des roches datant de −2,1 Ga à −600 Ma. Ce sont des ortho- et paragneiss datant de l'Icartien (−2,5 Ga à −1,6 Ga), des sédiments métamorphisés du Pentévrien (environ −1 Ga, on dit maintenant plutôt Mésoprotérozoïque), et des sédiments du Briovérien (Néoprotérozoïque) issus du démantèlement de la chaine cadomienne (Thomas, 2021 [29] ; Aubray et al., 2016 [2] ; Thomas, 2022 [30]). Ces affleurements sont tout de même assez rares et la question de la généralisation aux enveloppes de surface des informations apportées par ces affleurements se pose toujours.
La transition d'un océan ferrugineux vers un océan sulfuré au Paléoprotérozoïque
Pendant l'Archéen, les bassins océaniques profonds contenaient de grandes quantités de fer dissout, comme le montrent les dépôts massifs de BIF à cette époque. Le sulfure S2− est une espèce chimique qui réagit très rapidement avec le fer, et forme alors divers sulfures de fer comme la pyrite FeS2 ou la pyrrhotite, la mackinawite… Les dépôts de BIF dans l'océan archéen impliquent de faibles teneurs atmosphériques en oxygène (condition pour limiter les transferts d'oxygène vers les océans, donc pour avoir un océan profond anoxique) et de faibles teneurs océaniques en sulfures S2−. Ces faibles teneurs en sulfures impliquent de faibles apports en sulfates SO42−. En effet, les bactéries sulfato-réductrices (BSR) utilisent les sulfates comme accepteurs d'électrons (oxydants, comme l'oxygène pour la respiration aérobie) lors de la respiration et les réduisent en sulfures. Ces bactéries sont ubiquistes. Malgré leur métabolisme anaérobie, elles tolèrent de faibles teneurs en oxygène (contrairement aux Archées méthanogènes). À l'heure actuelle, il est très facile d'en isoler dans la zone à oxygène minimum océanique. Si les océans avaient reçu des fleuves une quantité importante de sulfates pendant l'Archéen, les océans profonds auraient été sulfureux comme la Mer Noire à l'heure actuelle. Les modèles récents suggèrent que les conditions sulfureuses de l'océan profond devinrent générales autour de 1,84 Ga, et le restèrent pendant tout le Mésoprotérozoïque et le début du Néoprotérozoïque (jusqu'à −580 Ma).
Cette transition a été étudiée dans les formations de Gunflint et de Rove (Poulton et al., 2004 [21]), dans l'Ontario (Canada). Ces formations consistent en des roches silico-clastiques déposées sur une plateforme connectée à l'océan profond (vu les quantités de fer nécessaires à la formation des BIF). La formation de Gunflint, datée à −1,878 Ga par des zircons contient des BIF, tandis que celle de Rove qui la surmonte, plus récente (avec sans doute un hiatus entre les deux) a été datée à −1,84 Ga.
Les auteurs se sont intéressés à la spéciation du fer, à savoir la forme chimique sous laquelle le fer se trouve dans ces formations. Le fer hautement réactif FeHR consiste en la phase pyrite (FeS2) et en toutes les phases susceptibles de former de la pyrite dans la colonne d'eau ou pendant la diagenèse précoce, à savoir la magnétite Fe3O4, les oxydes de fer FeOx et la sidérite FeCO3. Le fer restant (non réactif) est sous forme de silicates, peu réactifs. Le rapport entre le fer hautement réactif (FeHR) et le fer total FeT est un indicateur, ou proxy, des conditions de dépôt, oxiques ou anoxiques. À l'heure actuelle, dans les bassins anoxiques, le rapport FeHR/FeT est élevé, toujours supérieur à 0,38, ce rapport est très faible en conditions oxiques. La figure 1 présente l'évolution de la quantité totale de fer et de sa spéciation, ainsi que l'évolution des rapports isotopiques du soufre dans les formations de Gunflint et de Rove. La concentration du fer chute dans la formation de Rove, mais le rapport élevé FeHR/FeT indique que les conditions sont restées anoxiques. Dans le détail, le rapport FePyr/FeHR augmente très fortement dans la formation de Rove jusqu'à atteindre le niveau des bassins euxiniques (sulfurés) actuels.
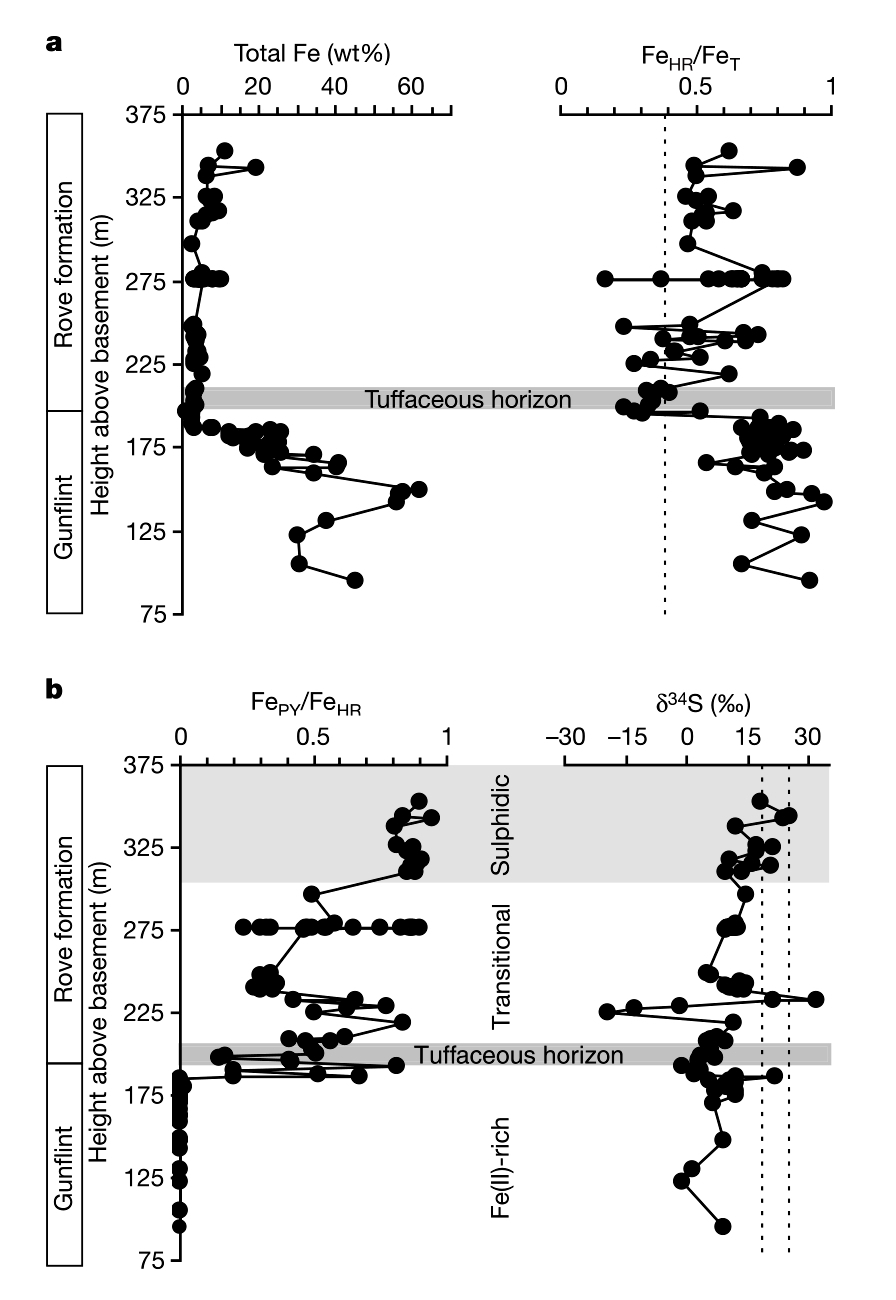
Source - © 2004 D’après Poulton et al. [21]
a. FeT and FeHR/FeT. La ligne pointillée représente un rapport de 0,38, en dessous duquel les conditions sont supposées oxygénées.
b. Fepy/FeHR et δ34S de la pyrite. Les lignes pointillées représentent une estimation des valeurs du δ34S des sulfates marins au début du Protérozoïque. Les bactéries réduisent donc une part majoritaire des sulfates marins issus de l’altération des continents d’où un faible fractionnement entre le soufre marin disponible et le soufre réduit (le fractionnement lié à l’activité bactérienne est d’autant moins marqué qu’une forte fraction du soufre initialement disponible est réduit – à l’extrême, si tout le soufre initialement disponible est réduit, ce soufre réduit à les mêmes rapports isotopiques que le soufre initial).
Des considérations physico-chimiques et biologiques simples permettent de comprendre cette évolution. L'augmentation de la teneur en oxygène atmosphérique à la fin du GOE a certainement augmenté les flux de sulfates vers les océans, puisque la pyrite détritique n'existait plus à ce moment (en oxydant la pyrite sur les continents, et en transportant les sulfates vers les océans via les fleuves). Les sulfates présents dans la colonne d'eau auraient été réduits en sulfures par des bactéries sulfato-réductrices, ces sulfures précipitant dans la colonne d'eau ou le sédiment pour former de la pyrite. La succession BIF puis argilites/sédiments siliceux pyritisés est caractéristique de la transition océan profond ferrugineux / océan profond sulfureux vers −1,8 Ga. La question de savoir si les océans étaient globalement sulfureux ou si ces conditions sulfureuses ne concernent que l'échelle locale n'est pas tranchée (Reinhard et al., 2013 [22]).
Ces conditions physico-chimiques océaniques vont durer un bon milliard d'années.
Des glaciations majeures à la fin du Néoprotérozoïque
Des affleurements canadiens et australiens (formation Elatina) prouvent l'occurrence de glaciations majeures à la fin du Néoprotérozoïque. Des dropstones de basses latitudes associées à des rythmites tidales attestent du caractère général de ces glaciations (Schmidt et al., 1991 [25]). Le déterminisme de ces glaciations est compris depuis peu, même si ses conséquences sur la biosphère restent discutées. Le supercontinent Rodinia se forme par accrétion de cratons continentaux il y a −1,3 à −1,1 Ga. Ce supercontinent se situe, d'après les enregistrements paléomagnétiques, entre les deux tropiques. Lors de l'éclatement de ce supercontinent, les petites plaques continentales restent en zone tropicale. Contrairement au Rodinia, plutôt aride au centre et arrosé en périphérie, ces plaques plus petites vont être beaucoup plus intensément arrosées, subir une érosion maximale, et l'altération des silicates calciques pompe le CO2 atmosphérique, ce qui produit l'englacement (Donnadieu et al., 2004 [9]). Attention, après cette chute, on estime la teneur en CO2 atmosphérique à 1000 ppm, à comparer aux 420 ppm actuels. Ce paradoxe est dû à la moindre activité du Soleil à l'époque. Ces glaciations semblent avoir eu une importance majeure pour l'émergence des eucaryotes modernes et des animaux (Brocks et al., 2017 [3] ; Brocks et al. 2023 [4]).
La transition vers un océan de nouveau ferrugineux puis oxygéné pendant le Néoprotérozoïque
Les formes de vie complexes sont apparues dans le registre sédimentaire vers −600 Ma. Ces formes sont représentées par les faunes d'Ediacara, des fossiles centimétriques à décimétriques, globalement distribués. Les faunes d'Ediacara se développent après la glaciation Gaskiers (−580 Ma) et disparaissent de manière abrupte au moment de l'explosion Cambrienne, 35 Ma plus tard. L'émergence des animaux à la fin du Néoprotérozoïque indique un épisode d'oxygénation des enveloppes externes à cette époque (Langlois, 2010 [16]). Les conditions de cette émergence sont encore discutées. Malgré une augmentation progressive de l'oxygénation des eaux de surface pendant le Mésoprotérozoïque, des BIFs apparaissent de nouveau au moment des grandes glaciations de la fin du Néoprotérozoïque. L'hypothèse des snowball Earths néoprotérozoïques (sturtienne, marinoenne et dans une moindre mesure Gaskiers) implique que la Terre était complètement recouverte d'une calotte de glace. Cette calotte surmontait un océan anoxique qui contenait suffisamment de fer (et suffisamment peu de sulfures) pour former les BIFs. Des affleurements exceptionnels au Canada, dans la péninsule d'Avalon (Newfoundland), permettent de préciser le scénario d'évolution de l'océan profond (Canfield et al., 2008 [6]). Les lits de dépôts de coulées de débris, les unités affaissées, la faible dispersion des directions de paléo-écoulements de courants de turbidité et l'absence de structures générées par les vagues indiquent que les sédiments ont été déposés sur une pente en eau profonde. Ces affleurements d'âge néoprotérozoïque représentent 15 Ma d'enregistrement. La séquence est surmontée de dépôts glaciaires et de carbonates de couverture datés à −580 Ma (épisode Gaskiers). La glaciation Gaskiers est la dernière glaciation majeure du Protérozoïque, représentée sur quatre continents (affleurements majeurs au Canada près de Boston, en Chine, en Norvège, en Écosse, en Irlande et en Australie) bien que ce soit un épisode moins sévère et global que les snowball Earths sturtienne et marinoenne. Des fossiles de type Ediacara sont fréquemment retrouvés après les dépôts Gaskiers.
L'étude de la spéciation du fer (forme chimique sous laquelle se trouve le fer) dans les dépôts néoprotérozoïques d'Avalon montre une abondance de carbonates et d'oxydes de fer sans sulfures : la colonne d'eau était donc ferrugineuse (le fer déposé est susceptible de réagir avec l'oxygène, car réduit) et non sulfureuse, ce qui représente une chimie océanique proche de celle des glaciations du Paléoprotérozoïque.
Dans les sédiments accumulés juste après la glaciation Gaskiers, et durant une période de 15 Ma, la spéciation du fer est comparable à celle des océans du Phanérozoïque, le fer ne réagit pas avec l'oxygène car il est entièrement oxydé. Les dépôts se sont donc effectués dans des conditions oxygénées. Les sédiments étant des sédiments de bas de pente et de plaine abyssale (turbidites, argiles des grands fonds), ces données prouvent l'oxygénation pérenne des eaux profondes à ce moment. Des organismes appartenant aux faunes édiacariennes vivent au fond de l'océan, ce qui corrobore son oxygénation. Le déterminisme de ces changements dans la chimie des océans ne fait pas consensus pour l'instant. La disparition des conditions “sulfureuses” serait due à la subduction accrue de sédiments sulfureux (contenant de la pyrite) dans le manteau, qui entrainerait un amoindrissement de la quantité de sulfures continentaux disponibles pour être altérés en sulfates (Canfield, 2004 [5]). L'augmentation de la teneur en oxygène post-Gaskiers serait due à des changements dans la nature des producteurs primaires.
En résumé. Le début du Protérozoïque est marqué par le Grand Évènement Oxydant (GOE, −2,45 à −2,2 Ga), pendant lequel se forment de volumineux gisements de fer sous forme de BIFs. L'atmosphère et l'océan superficiel deviennent partiellement oxygénés (entre 0,2 % et 8 % d'oxygène dans l'atmosphère) mais pas l'océan profond (Kump, 2008 [15]). À la fin de ces dépôts (vers −1,88 à −1,84 Ga), l'océan profond passe d'un état ferrugineux à un état sulfureux : l'oxygène de l'air (~2 %) permet l'oxydation des sulfures continentaux en sulfates, qui sont transportés par les fleuves jusqu'aux océans, réduits en sulfures par des bactéries dans l'océan profond et précipités avec le fer sous forme de pyrite. Ces conditions durent pendant un milliard d'années. À la fin du Protérozoïque, de grandes glaciations surviennent au cours de ce qu'on appelle le Cryogénien (−720 à −635 Ma), qui précède l'Édiacarien. Pendant ces grandes glaciations, l'océan profond redevient ferrugineux, de grandes quantités de sulfures ayant été enfouies dans le manteau par subduction (hypothèse considérée comme la plus probable actuellement) (Canfield, 2004 [5]). Après ces grandes glaciations (−580 Ma), l'océan profond est oxygéné, et les communautés édiacariennes sont fréquentes (Canfield et al., 2007 [7]).
La biosphère au Protérozoïque et ses relations avec les enveloppes externes
Les premiers eucaryotes retrouvés dans le registre fossile sont les Grypania spiralis, d'Empire Mine, au Nord du Michigan, à proximité de BIFs (Han et Runnegar, 1992 [13]). Ces fossiles datés de −2,1 Ga sont considérés comme étant ceux d'algues unicellulaires spiralées. Des fossiles pyritisés de métazoaires de cette époque sont retrouvés dans le bassin uranifère de Franceville, au Gabon. Si l'âge de la formation gabonaise n'est pas contesté (ce bassin qui a fourni gracieusement l'uranium nécessaire aux premières centrales nucléaires françaises a été intensément étudié, il a connu à Oklo une réaction en chaine naturelle), la nature métazoaire de ces fossiles n'est pas admise par tous les paléontologues (Langlois, 2010 [16]). On ne trouve pas de descendants de cette faune, il pourrait s'agir d'une expérience avortée de la vie. Vers −1,1 Ga, les premières preuves de colonisation des continents par les eucaryotes (Strother et al., 2011 [27]) apparaissent (affleurements du Torridonien du Nord de l'Écosse caractérisés par de superbes dunes éoliennes aux grains triés et émoussés (Lebeau et Ielpi, 2017 [18])).

Source - © 2017 D’après Lebeau et Ielpi [18]
Figure 2. Faciès éoliens du Torridonien d'Écosse
A et B : lame mince observée en lumière polarisée analysée. C : photographie d'un grès quartzeux. D : affleurement de dune éolienne avec stratifications entrecroisées et pendages élevés.
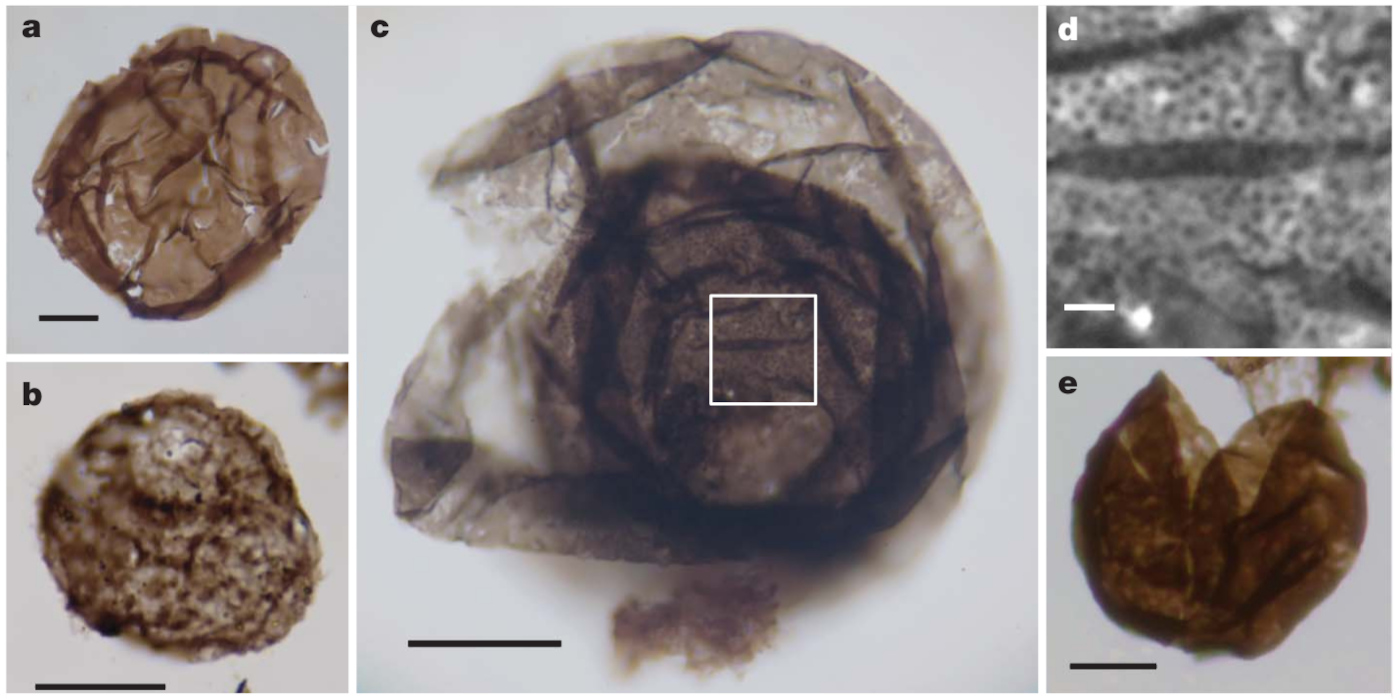
Source - © 2011 D’après Strother et al. [27]
Figure 3. Acritarches sphéromorphes du Torridonien d'Écossse (~1 Ga)
Barre d'échelle : 10 μm (a, b, c, e) ; 1 μm (d, détail de la figure c).
Une structure de déhiscence est supposée en d.
Plusieurs grandes questions se posent quant aux interactions biosphère-enveloppes externes pendant le Protérozoïque. Pour quelles raisons la teneur en oxygène varie-t-elle si peu (à l'exception des finalement modestes fluctuations mésoprotérozoïques) jusqu'au Cryogénien ? Pour quelles raisons la teneur en oxygène augmente-t-elle juste après la glaciation marinoenne ? Quel serait le lien entre les glaciations de la fin du Néoprotérozoïque et l'émergence des métazoaires ? On peut supposer un tel lien, car le froid intense est un facteur écologique qui pèse sur les êtres vivants, les glaciations ont très probablement entrainé la disparition d'eucaryotes inadaptés (Brocks et al., 2023 [4]) ; de plus, l'altération et l'érosion qui suivent une période glaciaire sont supposées apporter un flux important de nutriments vers l'océan, à l'origine d'évolutions rapides (Brocks et al., 2017 [3]).
Stase biologique et fixation de l'azote ?
Les géochimistes ont reconnu depuis longtemps le problème que pose la stase des teneurs en O2 entre le Grand Évènement Oxydant et l'oxygénation néoprotérozoïque. Une première explication consiste en l'apport continuel aux enveloppes externes de réducteurs comme le Fe(II) et le S2−, qui constituent des pièges à oxygène et des substrats pour les phototrophes anoxygéniques qui auraient ainsi dominé les phototrophes oxygéniques (et auraient été des compétiteurs pour le phosphore). Le second type d'explication repose sur la présence de facteurs limitants. Le molybdène, par exemple, est nécessaire aux cyanobactéries pour leur croissance, sa faible disponibilité à cause des conditions sulfureuses (le molybdène se liant très fortement aux sulfures) aurait limité la teneur en oxygène en restreignant la biomasse photosynthétique oxygénique. Avant l'apparition des photosynthèses, le dihydrogène H2 produit par des réactions géochimiques était le réducteur permettant la croissance autotrophe, à l'origine des ferrédoxines qui réduisaient le CO2. L'apparition de la photosynthèse anoxygénique avec un seul centre réactionnel a permis l'accès à des réducteurs plus abondants (Fe(II), sulfures) mais toujours limitants. La coopération entre deux systèmes photosynthétiques à l'origine de la photolyse de l'eau a changé la nature de la production primaire, en ce sens qu'il n'y avait plus de limite à cette production imposée par la faible disponibilité du donneur d'électrons (l'eau). Si l'abondance du donneur d'électrons n'est pas le facteur limitant, on peut proposer la faible disponibilité de l'azote.
Le carbone et l'azote entrent dans la biomasse à partir de réactions distinctes catalysées par des enzymes spécifiques. Les cyanobactéries fixent le carbone via le cycle de Calvin. La première étape est la carboxylation du ribulose 1,5 biphosphate, catalysée par la Rubisco. Bien que l'oxygène puisse entrer en compétition avec le CO2 pour cette enzyme, l'oxygène n'inactive pas la Rubisco. Pour l'azote, la situation est différente : il n'y a qu'une voie d'entrée pour l'azote dans la biosphère, via la nitrogénase. La nitrogénase catalyse la réduction du N2 atmosphérique ou dissout en ammonium, en utilisant des électrons qui proviennent d'une ferrédoxine réduite. Ces ferrédoxines sont également susceptibles de réduire l'oxygène. Toutes les nitrogénases actuellement connues partagent la même « enzyme ancestrale » et sont inactivées par l'oxygène (Esteves-Ferreira et al., 2017 [10]).
L'hypothèse de l'inhibition rétroactive des nitrogénases est une explication potentielle de la stase en oxygène (Allen et al., 2019 [1]). Les cyanobactéries doivent fixer du carbone mais aussi de l'azote. Il existe un seuil en oxygène, d'environ 2 % dans la phase gazeuse, à partir duquel les nitrogénases sont inactivées. Lorsque les cyanobactéries croissent, elles fixent carbone et azote, et libèrent de l'oxygène jusqu'au moment où la teneur en oxygène inhibe les nitrogénases. La figure 4 résume le mécanisme. Ce type d'inhibition rétroactive a été testé sur des cultures pures de cyanobactéries (Plectonema) en conditions contrôlées.

Source - © 2019 D’après Allen et al. [1]
Figure 4. Modèle d'inhibition rétroactive de la nitrogénase lors de la croissance des cyanobactéries
Lors de l'augmentation de la concentration en oxygène de la fin du Protérozoïque, diverses adaptations ont permis aux cyanobactéries de résoudre le problème posé par l'oxygène (différenciation des cellules akinètes, fixation de l'azote à l'obscurité…).
Essor des algues et oxygénation
Les horloges moléculaires placent l'origine des premières algues endosymbiotiques (Archaeplastidées qui comprennent les Chlorophytes, les Rhodophytes et les Glaucophytes) entre −1,9 Ga et −900 Ma (Sánchez-Baracaldo et al., 2017 [24]). Comme les algues laissent rarement un enregistrement sédimentaire indiscutable, on ne sait pas quand les Archaeplastidées ont commencé à jouer un rôle majeur dans les écosystèmes marins. Des informations complémentaires sont apportées par les biomarqueurs présents dans les membranes lipidiques. L'immense majorité des eucaryotes produit des stérols tétracycliques, qui régulent la physiologie membranaire, tandis que de nombreuses bactéries synthétisent des hopane-polyols pentacycliques. On connait de plus en plus de bactéries qui possèdent les gènes nécessaires à la synthèse des stérols. Stérols et hopane-polyols sont relativement stables vis-à-vis de la dégradation et sont conservés dans les strates sédimentaires sous forme de stéranes et d'hopanes (ou de dérivés stéroïdes / hopanoïdes). Bien que ces marqueurs possèdent également leurs propres biais et limites dans la précision des affiliations taxonomiques, leur étude permet d'estimer l'ordre de grandeur des abondances relatives des principaux groupes taxonomiques. Le rapport stéranes sur hopanes enregistre le flux relatif de lipides eucaryotes et procaryotes vers les sédiments, et donne une approximation relative de leurs biomasses respectives.
Des équipes ont compilé les données relatives aux enregistrements des rapports de ces lipides à la transition Protérozoïque-Phanérozoïque dans plusieurs formations sédimentaires de type schistes bitumineux (Brocks et al., 2017 [3]) : en Oman, dans le Bassin central Australien, dans le Grand Canyon (Arizona) et dans le lac Vättern (Suède). Les stéranes saturés les plus vieux apparaissent entre −820 et −720 Ma dans quatre bassins séparés du supercontinent Rodinia. Les rapports stéranes sur hopane (0,06 ± 0,10) sont largement en-dessous des rapports phanérozoïques (0,75 ± 1,1), suggérant un rôle mineur pour les eucaryotes. Dans le petit intervalle entre les snowball Earths sturtienne et marinoenne, entre −659 et −645 Ma, les stéroïdes deviennent plus abondants et plus variés. La domination des cyanobactéries aurait été abolie par des afflux de nutriments suite à la déglaciation du Sturtien. L'émergence des algues a sans doute créé des réseaux trophiques différents, conduisant à l'apparition d'organismes de plus en plus complexes. Il semble également que la taille de ces algues ait été à l'origine de l'oxygénation finale de la biosphère, à savoir l'Évènement d'Oxygénation Néoprotérozoïque (NOE) (Lenton et al., 2014 [19]). En effet, l’explication la plus traditionnelle est que l'augmentation de l'oxygénation de l'atmosphère a entrainé l'oxygénation de l'océan, conduisant à l'évolution des animaux. Mais une alternative a été proposée par Lenton et son équipe, pour qui l'évolution des eucaryotes, y compris des animaux, aurait oxygéné les océans sans requérir une augmentation de la teneur atmosphérique en oxygène ou de la production photosynthétique d'oxygène (Lenton et al., 2014 [19]). Les grands eucaryotes, abondants après le Cryogénien, auraient sédimenté plus rapidement à travers la colonne d'eau et réduit la consommation d'oxygène dans les eaux de surface (les cadavres n'auraient pas été décomposés / respirés). Le déclin de l'anoxie des eaux profondes aurait alors réduit le relargage de phosphore depuis les sédiments (les phosphates sont moins solubles dans des conditions oxydantes), réduisant à son tour la productivité globale et la demande en oxygène, et entrainant in fine l'oxygénation de ces océans. Cette oxygénation aurait alors permis l'évolution des macroorganismes (figure 5).

Source - © 2014 D’après Lenton et al. [19]
Deux phénomènes conduisent à la diminution du recyclage du carbone, donc à l'oxygénation : la grande taille des algues eucaryotes les fait chuter rapidement dans la colonne d'eau, empêchant leur “respiration” ; l'oxygénation des fonds marins empêche le recyclage du phosphore, et limite alors la productivité organique (détails dans le texte).
Glaciations fini-protérozoïques et diversification des eucaryotes
Les stérols des membranes cellulaires jouent un rôle fondamental dans leur stabilité. À basse température notamment, les stérols font office d'antigel et empêchent les membranes de se rigidifier. Comme tous les eucaryotes actuels possèdent des stérols, on suppose que LECA (Last Eukaryotic Common Ancestor, hypothétique ancêtre de tous les eucaryotes dont les caractères sont reconstruits par des méthodes phylogénétiques) en possédait. En se dégradant dans les sédiments, les stérols donnent des stéranes, qui sont dosés par les géochimistes. Les stéranes apparaissent dans le registre sédimentaire il y a 800 Ma, alors que les fossiles d'eucaryotes sont présents depuis 2,1 Ga (Grypania spiralis, Han et Runnegar, 1992 [13]). Cet écart était supposé dû à la dégradation des stéranes anciens.
L'équipe de Jochen Brocks (qui a démontré l'émergence des algues entre le Sturtien et le Marinoen) a fait l'hypothèse que les eucaryotes anciens possédaient des précurseurs de stérols mais pas de stérols, qui seraient apparus vers −800 Ma (Brocks et al., 2023 [4]). Ils ont d'abord réalisé une maturation artificielle d'un mélange de précurseurs des stérols, afin d'identifier leurs produits de dégradation. Ce sont ces molécules qu'ils ont recherchées (et trouvées) dans des schistes australiens datant de −1,64 Ga. Les protostéroïdes révèlent une biodiversité jusqu'ici inconnue d'eucaryotes et de bactéries produisant des protostérols abondants dans les environnements aquatiques de −1,64 Ga à −800 Ma. Les eucaryotes modernes apparaissent pendant le Tonien (ère qui précède le Cryogénien, à l'intérieur du Néoprotérozoïque), principalement sous forme d'algues rouges (Rhodophytes), vers −800 Ma. Ces eucaryotes se maintiennent après le Cryogénien, les autres eucaryotes ayant disparu, sans doute par incapacité à s’adapter au froid, selon la conclusion assez vraisemblable des auteurs.
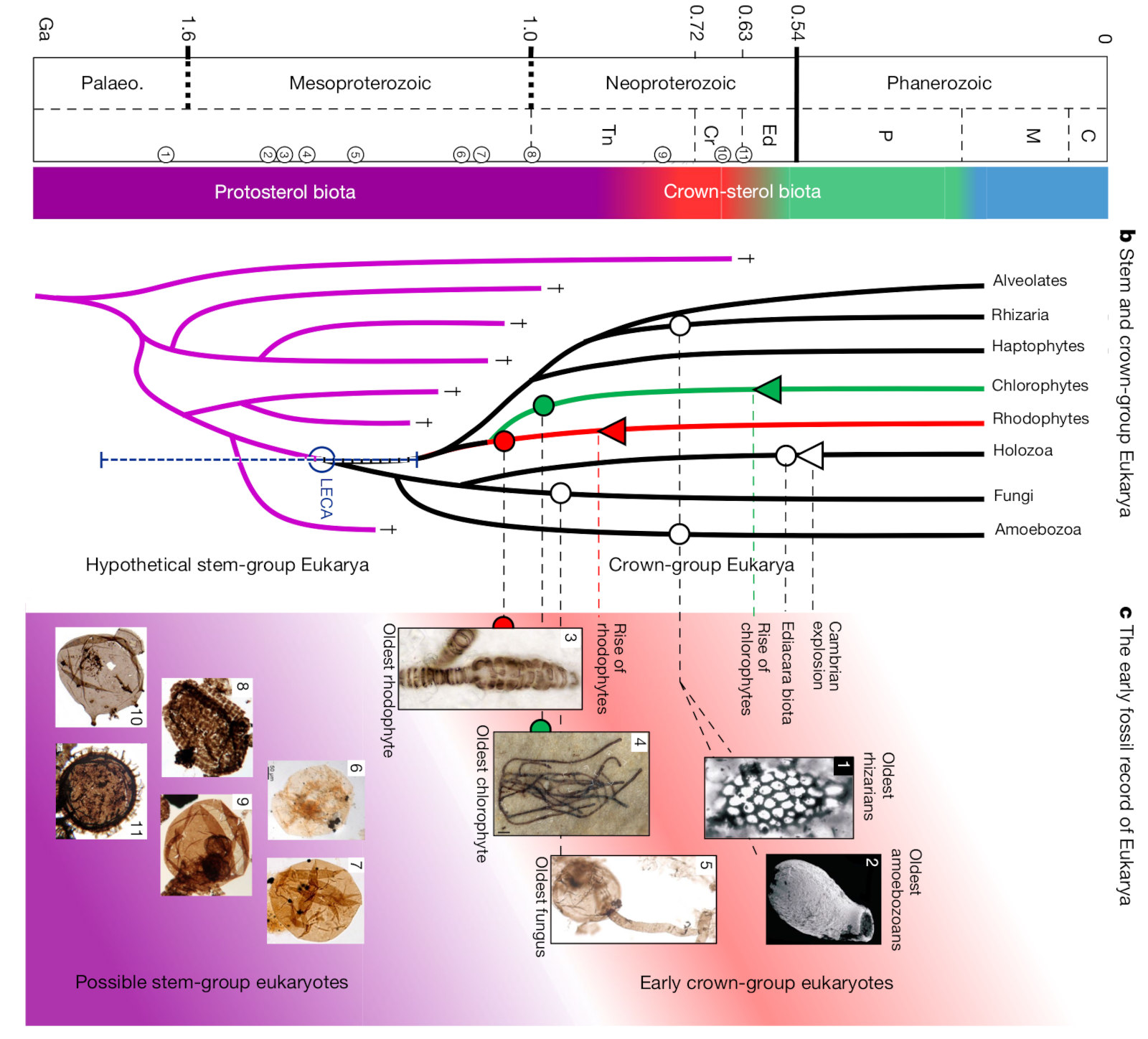
Source - © 2023 D’après Brocks et al. [4]
b. Arbre phylogénétique du domaine eucaryote. c. Microfossiles d'eucaryotes anciens (approximativement −1,1 à −0,7 Ga).
On observe au Protérozoïque, avant l'Édiacarien (−635 Ma), une longue stase biogéochimique, avec des teneurs atmosphériques en oxygène stables vers 2 % et des océans profonds anoxiques (sulfureux puis ferrugineux) (Canfield et al., 2008 [6]). Cette stase est peut-être liée à l'inhibition des nitrogénases cyanobactériennes par l'oxygène atmosphérique (Allen et al., 2019 [1]). Vers la fin du Protérozoïque, des épisodes de snowball Earths sont observés, liés à l'éclatement du supercontinent à de basses latitudes selon un axe Ouest-Est, qui a entrainé un pompage de CO2 par l'altération des silicates calciques (Donnadieu et al., 2004 [9]). La fin de la glaciation sturtienne entraine un important apport en nutriments à l'océan, qui aurait favorisé le développement des algues unicellulaires par rapport aux cyanobactéries (Brocks et al., 2017 [3]). Les algues sédimentant plus rapidement, leur carbone serait moins recyclé et l'oxygène aurait donc pu s'accumuler dans l'atmosphère. Une autre conséquence majeure des glaciations extrêmes du Cryogénien est la sélection des eucaryotes modernes, dont les membranes contiennent du cholestérol, les autres groupes d'eucaryotes n'ayant pas survécu au froid sans le rôle protecteur du cholestérol sur les membranes cellulaires (Brocks et al., 2023 [4]).
Bibliographie
J.F. Allen, B. Thake, W.F. Martin, 2019. Nitrogenase Inhibition Limited Oxygenation of Earth, Trends Plant Sci., 24, 1022-1031 [Open Access]
A. Aubray, C. Langlois, D. Mollex, P. Thomas, 2016. Les orthogneiss de Port-Béni (Côtes d'Armor) : un affleurement des roches les plus anciennes de France métropolitaine, Planet Terre - ISSN 2552-9250 [Accès libre]
JJ. Brocks, A.J.M. Jarrett, E. Sirantoine, C. Hallmann, C.Y. Hoshino, T. Liyanage, 2017. The rise of algae in Cryogenian oceans and the emergence of animals, Nature 548, 578-581 [pdf]
J.J. Brocks, B.J. Nettersheim, P. Adam, P. Schaeffer, A.J.M. Jarrett, N. Güneli, T. Liyanage, L.M. van Maldegem, C. Hallmann, J.M. Hope, 2023. Lost world of complex life and the late rise of the eukaryotic crown, Nature, 618, 7966, 767-773 [pdf]
D.E. Canfield, 2004. The evolution of the Earth surface sulfur reservoir, American Journal of Science, 304, 10, 839-861 [Free Access]
D.E. Canfield, S.W. Poulton, A.H. Knoll, G.M. Narbonne, G. Ross, T. Goldberg, H. Strauss, 2008. Ferruginous conditions dominated later neoproterozoic deep-water chemistry, Science, 321, 5891, 949-952 [pdf]
D.E. Canfield, S.W. Poulton, G.M. Narbonne, 2007. Late-Neoproterozoic Deep-Ocean Oxygenation and the Rise of Animal Life, Science, 315, 5808, 92-95 [pdf]
M.E. Clapham, J.A. Karr, 2012. Environmental and biotic controls on the evolutionary history of insect body size, Proc. Natl. Acad. Sci. U.S.A., 109, 27, 10927-1093 [Free Access]
Y. Donnadieu, Y. Goddéris, G. Ramstein, A. Nédélec, J. Meert, 2004. A 'snowball Earth' climate triggered by continental break-up through changes in runoff, Nature, 428, 303-306 [pdf]
A.A. Esteves-Ferreira, J.H.F. Cavalcanti, M.G.M.V. Vaz, L.V. Alvarenga, A. Nunes-Nesi, W.L. Araújo, 2017. Cyanobacterial nitrogenases: phylogenetic diversity, regulation and functional predictions, Genet Mol Biol, 40, 261-275 [CC BY 4.0]
J. Farquhar, H. Bao, M.H. Thiemens, 2000. Atmospheric influence of Earth's earliest sulfur cycle, Science 289, 5480, 756-758
R. Frei, C. Gaucher, S.W. Poulton, D.E. Canfield, 2009. Fluctuations in Precambrian atmospheric oxygenation recorded by chromium isotopes, Nature, 461, 250-253 [pdf]
T.M. Han, B. Runnegar, 1992. Megascopic Eukaryotic Algae from the 2.1-Billion-Year-Old Negaunee Iron-Formation, Michigan, Science 257, 5067, 232-235
D.J. Horne, S.N. Brandão, I.J. Slipper, 2011. The Platycopid Signal deciphered: Responses of ostracod taxa to environmental change during the Cenomanian–Turonian Boundary Event (Late Cretaceous) in SE England, Palaeogeography, Palaeoclimatology, Palaeoecology, 308, 3-4, 304-312 [pdf]
L.R. Kump, 2008. The rise of atmospheric oxygen, Nature, 451, 277-278 [pdf]
C. Langlois, 2010. De Burgess à Franceville (Gabon) : les plus anciennes traces fossiles de pluricellulaires, Planet-Terre, ISSN 2552-9250 [Accès libre]
C. Langlois, 2018. Les « plumes » de l'Édiacarien, un groupe animal disparu ?, Planet-Terre, ISSN 2552-9250 [Accès libre]
L.E. Lebeau, A. Ielpi, 2017. Fluvial channel-belts, floodbasins, and aeolian ergs in the Precambrian Meall Dearg Formation (Torridonian of Scotland): Inferring climate regimes from pre-vegetation clastic rock records, Sedimentary Geology, 357, 53-71
T.M. Lenton, R.A. Boyle, S.W. Poulton, G.A. Shields-Zhou, N.J. Butterfield, 2014. Co-evolution of eukaryotes and ocean oxygenation in the Neoproterozoic era, Nature Geoscience, 7, 257- 265 [pdf]
H. Martin, J.-F. Moyen, R. Rapp, 2010. The sanukitoid series: magmatism at the Archaean–Proterozoic transition, in Sixth Hutton Symposium on The Origin of Granites and Related Rocks: Proceedings of a Symposium held in Stellenbosch, South Africa, 2- 6 July 2007, J.D. Clemens, C. Donaldson, C.D. Frost, A.F.M. Kisters, J.-F. Moyen, T. Rushmer, G. Stevens (eds), Geological Society of America
S.W. Poulton, P.W. Fralick, D.E. Canfield, 2004. The transition to a sulphidic ocean approximately ~1.84 billion, Nature, 431, 173-177
C.T. Reinhard, N.J. Planavsky, L.J. Robbins, C.A. Partin, B.C. Gill, S.V. Lalonde, A. Bekker, K.O. Konhauser, T.W. Lyons, 2013. Proterozoic ocean redox and biogeochemical stasis, Proc. Natl. Acad. Sci. U.S.A., 110, 5357-5362 [Free Access]
R. Rye, H.D. Holland, 1998. Paleosols and the evolution of atmospheric oxygen: a critical review, American Journal of Science, 298, 8, 621-672 [Free Access]
P. Sánchez-Baracaldo, J.A. Raven, D. Pisani, A.H. Knoll, 2017. Early photosynthetic eukaryotes inhabited low-salinity habitats, Proc. Natl. Acad. Sci. U.S.A., 114, 37, E7737-E7745 [Free Access]
P.W. Schmidt, G.E. Williams, B.J.J. Embleton, 1991. Low palaeolatitude of Late Proterozoic glaciation, Earth and Planetary Science Letters, 105, 355-367
S.C. Spinks, E.A. Sperling, R.L. Thorne, F. LaFountain, A.J.R. White, J. Armstrong, M. Woltering, I.M. Tyler, 2023. Mesoproterozoic surface oxygenation accompanied major sedimentary manganese deposition at 1.4 and 1.1 Ga, Geobiology, 21, 28-43 [Open Access]
P.K. Strother, L. Battison, M.D. Brasier, C.H. Wellman, 2011. Earth's earliest non-marine eukaryotes, Nature, 473, 505-509 [pdf]
P. Thomas, 2011. Les galets de pyrite de l'Archéen du Witwatersrand (Afrique du Sud) et la teneur en dioxygène de l'atmosphère, Planet-Terre, ISSN 2552-9250 [Accès libre]
P. Thomas, 2021. Les migmatites et le métamorphisme panafricains (cadomiens) de la région de Saint-Malo (Ille et Vilaine), Planet-Terre, ISSN 2552-9250 [Accès libre]
P. Thomas, 2022. Se promener sur la Pointe du Grouin (Cancale, Ille et Vilaine) : granite schistosé, courants de marée…, Planet-Terre, ISSN 2552-9250 [Accès libre]
S. Xiao, M. Laflamme, 2009. On the eve of animal radiation: phylogeny, ecology and evolution of the Ediacara biota, Trends in Ecology & Evolution, 24, 1, 31-40
S. Xiao, X. Yuan, A.H. Knoll, 2000. Eumetazoan fossils in terminal Proterozoic phosphorites?, Proc. Natl. Acad. Sci. U.S.A., 97, 25, 13684-13689 [Free Access]
[1] Un Δ est, ici, une différence de δ pondérée de sorte que la différence soit nulle en cas de “respect” de la dépendance à la masse. Ainsi, par exemple, Δ33S = δ33S − 0,515 × δ34S car la différence de masse 33S-32S est environ la moitié (0,515 exactement) de la différence de masse 34S-32S.




