
Article | 15/03/2007
L'Évolution montrée par les fossiles
15/03/2007
Résumé
Explications et données pour réfuter l'affirmation selon laquelle pas une seule des "formes transitoires" dont les évolutionnistes mettaient en avant qu'il devait en exister des millions n'a été trouvée.
Table des matières
L'argumentaire anti-évolutionniste
L'affirmation
« Le darwinisme ne met plus en avant les fossiles comme preuves de l'évolution. Car dans toutes les fouilles menées depuis le 19ème siècle, pas une seule des « formes transitoires », dont les évolutionnistes mettaient en avant qu'il devait en exister des millions, n'a été trouvée. »
Détournements et falsifications du raisonnement évolutionniste
Une telle phrase est susceptible de troubler les élèves par le fait que :
- elle emploie le même vocabulaire que celui de l'enseignant à propos de l'Évolution : forme transitoire ou « chaînon manquant », fossiles, etc ;
- elle fait appel à des conceptions obsolètes mais répandues : les idées d'un « progrès évolutif », d'une « chaîne des êtres » et d'une évolution graduelle et très lente ;
- elle dévoie des explications correctes quant au statut des espèces fossiles par rapport aux espèces actuelles et à leur contribution à la compréhension des processus évolutifs.
L'exploitation de conceptions populaires erronées concernant l'Évolution
L'évolution ne procède pas selon le schéma d'une modification progressive d'un organisme vers un autre, schéma qui s'associe généralement à l'idée d'un « progrès évolutif ». Cette vision reste malheureusement très enracinée dans le grand public, comme en témoignent les multiples variantes de la fameuse image de « l'évolution humaine » (figure ci-dessous).
La conception de l'évolution que suggère ce genre d'illustration (figure ci-dessous) est fondamentalement fausse.
- Rien ne permet actuellement d'affirmer que l'ancêtre commun de l'Homme et du Chimpanzé était morphologiquement semblable au Chimpanzé actuel. Rien n'indique non plus que le Chimpanzé en tant qu'espèce ait préexisté à l'espèce humaine ou aux autres espèces des genres Australopithecus et Homo.
- Sauf dans le cas d'évolution anagénétique (modification progressive d'une espèce), une forme n'en remplace pas une autre. Australopithèques et représentants du genre Homo ont coexistés pendant plusieurs millions d'années, plusieurs espèces du genre Homo ont également coexisté.
- les « tendances évolutives », par exemple l'augmentation de taille des espèces souvent constatée chez les grands groupes de Mammifères au cours de leur évolution au Tertiaire, peuvent s'inverser et ne sont pas systématiques (avec les cas fameux du nanisme (et du gigantisme) insulaire (figure ci-après), bien documentés chez les Mammifères et dont l'Homo floresiensis serait un exemple d'autant plus frappant qu'il touche notre propre groupe, le genre Homo).

Source - © 2000 Pour la Science / C. Restier (MNHN)
Figure 1. Éléphant nain de Sicile
Exemple du phénomène de nanisme insulaire chez les grands mammifères.
Malgré cela, cette iconographie reste abondamment employée, même et y compris par les partisans de l'évolution sous forme d'images humoristiques conçues pour combattre le message anti-évolutionniste.

Source - © 2007 Cyril Langlois
Figure 2. Une publicité chinoise pour une société de télécommunications. Kunming, Yunnan, Chine
Une déclinaison parmi tant d'autres de la représentation erronée de l'évolution humaine.
Rappelons que l'évolution n'est pas forcément graduelle ni lente (à l'échelle des temps géologiques), même si, effectivement, des cas de changement progressif de toute une population sont documentés dans le registre fossile, en particulier chez les micro-organismes planctoniques (évolution par anagenèse ou évolution anagénétique) : par exemple la taille moyenne de la population augmente, l'ornementation d'un test se modifie. Dans ce cas, c'est l'espèce dans son ensemble qui change. Rigoureusement, c'est toujours la même espèce, même si l'on peut décider finalement de désigner sous deux noms d'espèces différentes les formes les plus anciennes et les formes les plus récentes, pour mieux dater les sédiments qui contiennent ces formes.
L'interprétation évolutionniste
Les fossiles peuvent montrer l'apparition de nouvelles espèces
Contrairement à l'affirmation créationniste, les fossiles peuvent montrer comment apparaissent de nouvelles espèces. La figure ci-après en est un exemple. Les études les plus claires concernent souvent des micro-organismes planctoniques (foraminifères, radiolaires, ostracodes, etc.) car ces organismes ont l'avantage d'être abondant dans le registre fossile. Ils fournissent par conséquent des populations statistiquement significatives et une image relativement représentative de la variabilité des populations à un moment donné. De plus, les sédiments marins constituent des enregistrements relativement continus et assez longs, avec parfois une excellente résolution temporelle. En contrepartie, ces organismes et les enseignements qu'ils nous procurent restent inconnus du grand public.
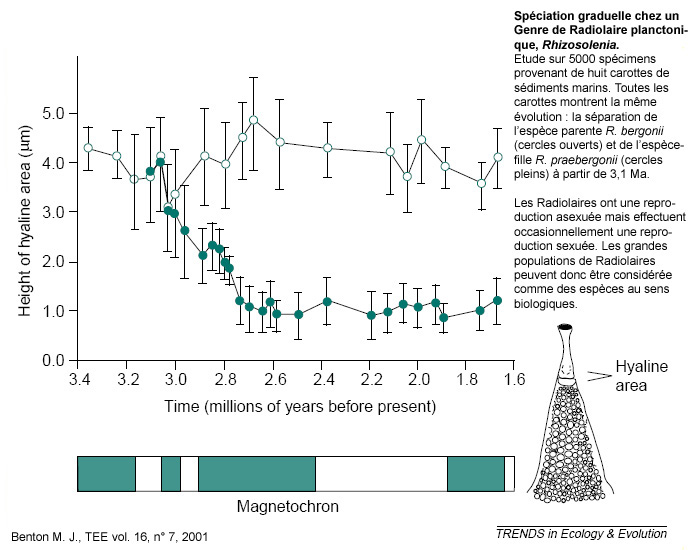
Source - © 2001 M.J. Benton, TEE, vol. 16, n°7
Deux populations, différentiables par la hauteur de l'aire hyaline, au sommet du test siliceux, se différencient à partir d'une espèce initiale. La bonne résolution temporelle de l'enregistrement fossile permet de voir la progression de cette séparation. En ce sens, les radiolaires retrouvés entre 3,0 et 2,8 Ma peuvent être qualifié de « formes de transition ».
Si l'on ne dispose pas de cas aussi clairs pour les organismes plus grands, c'est essentiellement en raison de la rareté des fossiles macroscopiques par rapports aux fossiles microscopiques : les organismes macroscopiques sont fondamentalement moins abondants que les microorganismes, leurs milieux de vie sont, pour beaucoup, moins propices à la fossilisation, etc. C'est l'une des raisons pour lesquelles, en général, l'ancêtre commun de deux espèces est présenté comme « virtuel ».
Par ailleurs, chez les « grands » organismes pluricellulaires, il semble que les évolutions enregistrées par les fossiles suivent souvent le schéma évolutif des « équilibres ponctués », c'est-à-dire qu'une espèce resterait morphologiquement stable sur de longues durées (des milliers ou millions d'années) et donnerait naissance à de nouvelles espèces, elles aussi stables, par des événements de spéciation très brefs à l'échelle des temps géologiques (centaines, milliers ou dizaines de milliers d'années). Le déroulement de ces spéciations est donc invisible dans le registre fossile, ce qui ne signifie pas, au contraire, que les mécanismes de la spéciation soient différents de ceux définis à partir des organismes actuels. Il pourrait s'agir, par exemple, de spéciation allopatrique, par isolement d'une petite population de l'espèce ancestrale et dérive génétique.
Dans ces cas-là, en effet, « les fossiles ne montrent pas les mécanismes de l'évolution », mais ils montrent néanmoins clairement le phénomène d'évolution, l'espèce « fille », descendante, pouvant sans ambiguïté être rapprochée de l'espèce « mère », ancestrale, dont elle est issue. Les fossiles montrent donc bien l'apparition de nouvelles espèces par évolution.
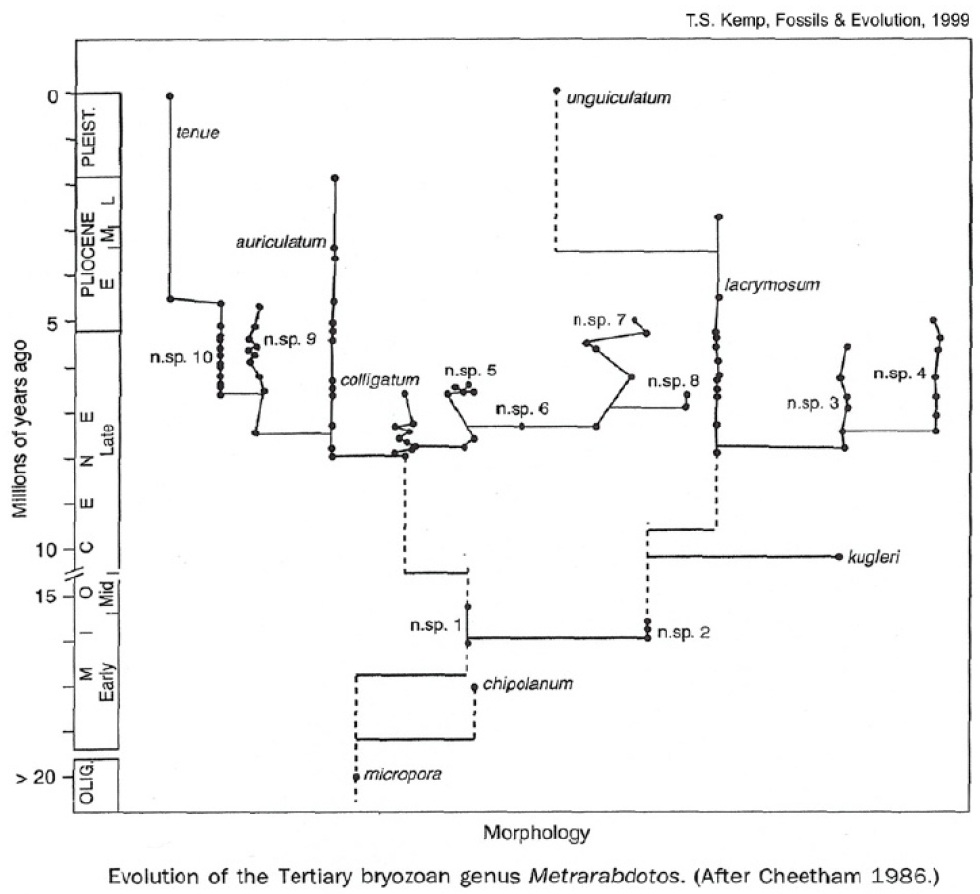
Source - © 2001 M.J. Benton, TEE, vol. 16, n°7
Figure 4. Évolution selon le modèle des "équilibres ponctués" pour le genre de Bryozoaires Metrarabdotos
L'apparition de nouvelles formes (espèces paléontologiques) s'effectue « brutalement » à l'échelle des temps géologiques et les nouvelles espèces restent ensuite stables pendant de longues durées, mais chaque forme nouvelle peut être rapprochée d'une espèce parente dont elle serait issue par spéciation.
Il n'y a pas de « chaîne des êtres », mais des buissons évolutifs
Les données paléontologiques montrent souvent que l'apparition d'un nouveau groupe d'organismes, ou d'une nouvelle espèce, s'effectue par scission d'une population ou d'un groupe en deux sous-ensembles, pouvant ensuite évoluer indépendamment et se scinder à leur tour. Partant d'une espèce initiale, un groupe pourra se diversifier en plusieurs espèces (si celles-ci peuvent occuper une niche écologique, c'est-à-dire soit être compétitives par rapport aux autres organismes qui occupent cette niche, soit présenter des caractéristiques particulières qui leur permettent d'occuper une position écologique vacante). Au cours de cette diversification, une nouvelle espèce peut coexister (dans le temps et dans l'espace) avec l'espèce dont elle dérive (cf. les figures ci-dessus) et avec les autres espèces nouvellement apparues dans le groupe.
De même, une espèce « dérivée » peut fort bien disparaître — quelle que soit la cause de son extinction — avant l'espèce « archaïque » dont elle s'est différenciée, un phénomène que l'idée de « progrès évolutif » rejette à priori (cf. sur la figure précédente le cas de l'espèce n. sp. 9 et de son ancêtre M. auriculatum).
« Transition évolutive » ne signifie pas « fonctionnellement transitoire »
L'argument anti-évolutionniste amalgame deux significations de la notion de « transition évolutive ».
- Dans la conception évolutionniste de cette notion, un organisme peut présenter des caractères qui, a posteriori, apparaissent comme des transitions entre les caractéristiques d'organismes plus anciens et plus récents. Chez tous ces organismes, les caractères considérés sont fonctionnels, au sens où ils assurent une fonction avec une efficacité suffisante pour permettre la survie de l'organisme.
- La nageoire des poissons sarcoptérygiens, sans doigts, est efficace pour sa fonction, la nage. Le membre chiridien des tétrapodes terrestres archaïques est suffisamment efficace pour permettre à l'animal une locomotion terrestre. Le membre du tétrapode primitif Tiktaalik rosae, sans doigts mais que l'animal peut poser sur le sol pour soulever sa tête hors de l'eau, était probablement efficace pour stabiliser l'animal dans un cours d'eau peu profond et lui permettre de chasser à l'affût, à l'image des crocodiles actuels. A posteriori, le membre de Tiktaalik rosae apparaît comme une « transition » entre nageoire sarcoptérygienne et chiridium tétrapode. Mais en réalité, chacun de ces trois animaux est fonctionnel et adapté à un environnement et un mode de vie qui lui est propre.
- Autre exemple : le site Understanding evolution, de l'Université de Berkeley, propose comme illustration d'une transition le cas des cétacés, en montrant les crânes de deux cétacés fossiles, Pakicetus et Aetiocetus, et d'un cétacé actuel, le Beluga. Pakicetus, le plus ancien représentant connu des cétacés (datés d'environ 50 Ma) était un quadrupède surtout terrestre, dont le mode de vie était peut-être proche de celui du raton-laveur. Chez cet animal, les narines s'ouvrent à l'extrémité du museau. Chez Aetiocetus, plus récent de 25 Ma environ et déjà pleinement aquatique, les narines se situaient au centre du museau, alors qu'elles s'ouvrent, chez le Béluga comme chez les autres baleines actuelles, au sommet du crâne. Dans ce cas-ci, on peut considérer cette transition comme une adaptation progressive et de plus en plus perfectionnée à la vie en milieu aquatique (l'animal n'a plus besoin d'émerger la tête de l'eau pour respirer), et Aetiocetus est bien un fossile intermédiaire dans ce processus d'adaptation. Pour autant, même s'il n'était pas aussi perfectionné que le Béluga actuel, Aetiocetus, avec ses narines en position médiane sur le museau, était certainement un animal pélagique tout à fait efficace et concurrentiel par rapport aux autres animaux avec lesquels il pouvait se trouver en compétition.
- Dans le discours anti-évolutionniste, l'idée qu'un organisme représente une « transition évolutive » entre deux états connus (par exemple « poisson » et « tétrapode ») signifie forcément que cet organisme n'est qu'un « juste milieu » entre deux pôles. Un tel animal hybride, à mi-chemin entre deux « états stables », chacun adaptés à des environnements ou à des modes de vie différents, est forcément une chimère. Il est très improbable qu'un tel organisme puisse apparaître au cours de l'évolution sauf à envisager l'existence d'un environnement lui aussi « intermédiaire » auquel il serait adapté. Cette possibilité est évidemment plus improbable encore si tous les caractères morpho-anatomiques et physiologiques de cet organisme hypothétique devaient eux aussi être des « intermédiaires ».

Source - © 2007 University of California, Museum of Paleontology
L'argumentaire anti-évolutionniste confond donc sciemment cette notion d'organisme « chimère » avec celle d'un organisme montrant des caractères particuliers qui, a posteriori, apparaissent comme des étapes dans un processus évolutif. On remarquera également que supposer qu'un organisme intermédiaire, une « chimère », puisse exister, relève d'une vision finaliste et dirigée de l'évolution : on ne peut concevoir une telle forme « transitionnelle » que si l'on connaît déjà, ou que l'on peut prévoir, les deux pôles entre lesquels cette transition vient s'insérer. L'évolution étant un processus historique, l'aspect d'un organisme ou d'un caractère dérivé n'est pas prévisible à l'avance. Un organisme n'évolue jamais « vers » une forme future, supposée « meilleure » ou plus perfectionnée. En ce sens, et en renversement de l'argument anti-évolutionniste, le fait que les fossiles ne soient jamais de parfaites « transitions », mais plutôt des mosaïques de caractères « ancestraux » et « dérivés » est parfaitement en accord avec l'idée d'évolution darwinienne.
Phylogénie cladistique et virtualité des ancêtres communs
Une dernière notion que déforme l'affirmation créationniste mentionnée plus haut est celle du caractère virtuel de l'ancêtre commun dans les phylogénies cladistiques, une notion qu'il n'est pas facile de faire passer aux élèves et aux étudiants.
Il faut rappeler ici que les phylogénies cladistiques (ou autres) sont des représentations sous forme d'arbres hiérarchiques des relations de parentés entre les taxons étudiés. Même si elles sous-tendent effectivement une hypothèse sur l'histoire évolutive du groupe étudié et des taxons qu'il contient, elle ne représente pas ce qui s'est réellement passé : une expérience de laboratoire dans laquelle un nouveau taxon de drosophiles ou de bactéries ferait son apparition par spéciation à partir d'un taxon préexistant conduira aussi à un arbre phylogénétique dans lequel les deux taxons, « l'ancestral » et le « dérivé » seront représentés au bout d'une branche et relié par une fourche dont le nœud avec le groupe externe (un autre taxon de drosophile ou de bactérie), correspondant à « l'ancêtre commun », sera virtuel. En réalité, la relation ancêtre-descendant est parfaitement constatée au cours de l'expérience, c'est la représentation phylogénétique qui introduit la « virtualité de l'ancêtre ».
Dans le cas des fossiles, ou des études cladistiques sur des taxons de haut niveau (Genres, Familles, Classes entières), la situation est encore compliquée par le fait que chaque branche de l'arbre représente fréquemment tout un groupe monophylétique de rang supérieur à l'espèce (telle famille de trilobites, tel genre de mammifères, etc.), mais que ce groupe n'est représenté, en général, que par une seule espèce voire un seul fossile.
Jouer ainsi sur l'idée d'inexistence des ancêtres communs (donc des « transitions ») constitue donc un détournement complet des objectifs et des résultats de l'analyse phylogénétique et une mise à profit scandaleuse de la difficulté conceptuelle entourant ce mode de raisonnement.
Conclusion
Cette attaque créationniste de la théorie de l'évolution qui affirme qu'il n'existe pas de « fossiles de transition » alors qu'ils seraient nécessaires pour démontrer la réalité de l'évolution est donc fausse : le déroulement d'un phénomène de spéciation, c'est-à-dire l'apparition d'une ou plusieurs nouvelles formes à partir d'une population préexistante et en passant par des formes qui sont forcément « de transition » peut être détaillé grâce au registre fossile, même si ce cas reste rare en raison des dégradations liés aux processus de fossilisation.
Plus largement, le registre fossile peut présenter des organismes dont certains états de caractères peuvent, a posteriori, apparaître comme des états de transition entre deux autres états connus. Mais il convient de ne pas confondre cet aspect transitoire dans la forme d'un organe ou d'une structure avec un fonctionnement transitoire ou intermédiaire, donc inopérant, de cet organe.
Références bibliographiques utiles
- Mark Ridley, 1998. Évolution biologique, De Boek Université.
- Francine Brondex, 1999. Évolution : synthèse des faits et théorie, Dunod
- Understanding evolution : un site web particulièrement bien illustré, spécifiquement destiné à expliquer à un public d'élèves du secondaire les principes de la théorie de l'évolution et les multiples arguments qui la soutiennent. Seul obstacle pour les élèves francophones : la langue anglo-saxonne.



