
Article | 15/05/2002
Arguments paléontologiques en faveur de l'évolution
15/05/2002
Résumé
Apport de la paléontologie à la compréhension de la théorie de l'évolution.
Table des matières
Introduction
L'évolution des êtres vivants, ou "Évolution", est définie comme l'ensemble des processus par lesquels les organismes se transforment, dans leur structure et leur comportement, au fil des générations, d'ancêtre à descendants.
Elle constitue actuellement une notion centrale des sciences biologiques. Largement acceptée dans son principe en Europe, elle reste fortement combattue, pour des raisons religieuses, en Amérique du Nord, en dépit des nombreuses données qui la soutiennent. Outre les preuves obtenues par les expériences de sélection artificielle menées par les biologistes, bon nombre de ces données découlent des travaux paléontologiques et géologiques.
Rappelons d'abord que la géologie, avec Buffon (d'après l'épaisseur des sédiments et le principe de l'actualisme), Lamarck puis Lyell, ont établi le grand âge de la Terre, condition préalable pour concevoir les processus évolutifs, supposés lents à l'échelle humaine. Quel âge a la Terre ?
 Figure 1. Le comte de Buffon (1707-1788) |  Figure 2. Lamarck (1744-1829) |  Figure 3. Charles Darwin (1809-1882) |
Ces mêmes chercheurs se sont également intéressés à la paléontologie, qui, elle, cherche et décrit les formes anciennes et disparues de la vie.
Nous expliquerons d'abord comment les données qu'elle fournit ont contribué, depuis le XIXème siècle, à l'émergence de l'idée d'évolution des êtres vivants, que les observations biologiques sur l'actuel de Darwin viendront étayer en 1859. Après quoi, nous verrons sur quelques cas comment cette discipline peut indiquer des modalités d'évolution, et proposer des mécanismes théoriques de ce phénomène.
Comment la paléontologie montre l'évolution
Paléontologie et histoire du vivant
La paléontologie permet avant tout d'établir que des êtres vivants différents ont existé au cours des temps géologiques, donc de décrire une histoire. Cela n'implique pas l'Évolution puisque Cuvier avait constaté que les faunes fossiles différaient des faunes actuelles, mais attribuait cela à des extinctions et créations successives d'espèces, qui restaient ensuite immuables jusqu'à l'extinction suivante. De même, il avait montré l'existence de grands plans d'organisation animaux (Vertébrés, Mollusques, Articulés, Zoophytes, selon sa classification), qu'il supposaient issus de créations séparées.

Figure 4. Le baron Cuvier (1769-1832)
Cependant, l'ordre d'apparition maintenant établi pour les différentes formes vivantes prouve déjà que les formes les plus complexes sont aussi les plus récentes (Lethiers, Fig.22). Ce constat invalide l'idée d'une création divine initiale des organismes.
Lamarck, lui, avait déjà noté que les faunes de mollusques fossiles différaient d'autant plus des faunes actuelles qu'elles étaient anciennes, et en avait tiré l'idée de modifications au cours du temps à partir d'un état primitif simple apparu par génération spontanée. De telles créatures simples apparaîtraient continûment pour s'engager alors dans un processus de complexification progressive (la « montée dans l'échelle des êtres »).
Or, on constate que l'état unicellulaire est resté la seule forme vivante pendant plusieurs milliards d'années (Lethiers, Fig.20), alors que les épisodes de grandes extinctions ont été suivi quasi aussitôt d'un renouvellement des faunes et/ou flores sans retour pour autant à un état de complexité inférieur, ce qui rend improbable l'hypothèse de créations successives des organismes par génération spontanée. Néanmoins, l'idée d'évolution apparaît déjà avec Lamarck, même si le mécanisme qu'il imagine, l'hérédité des caractères acquis, s'est avéré faux avec la découverte de la génétique mendélienne. Reste l'idée du lien généalogique des êtres vivants (au moins à l'intérieur des grands "types"). Là encore, la paléontologie va apporter des arguments.
Paléontologie et notion d'homologie
La paléontologie montre que les organismes anciens, au moins depuis l'apparition des êtres multicellulaires, et comme les actuels, présentent entre eux des structures ressemblantes, plus ou moins marquées, dites homologies. Ces homologies ont plusieurs caractéristiques.
Elles permettent, la plupart du temps, de classer les organismes anciens selon l'organisation hiérarchique définie par Linné.
Elles ne sont pas explicables par des nécessités fonctionnelles : l'exemple simple ici est la pentadactylie des tétrapodes.
Les fossiles des premiers tétrapodes, comme Acanthostega (Dévonien), présentaient un membre à plus de 5 doigts (8).
 Source - © 1997 La Recherche n°296, p.59 |  Source - © 1997 La Recherche n°296, p.59 Figure 6. Reconstruction d'Acanthostega |
La rétention chez tous les tétrapodes qui suivent, d'un membre à 5 doigts s'explique aisément par l'hypothèse d'un lien de parenté entre eux, et par leur origine dans une forme proche d'Acanthostega , mais pentadactyle.
Ces homologies peuvent être altérées et moins visibles chez certaines formes, souvent spécialisées, mais que l'anatomie comparée (pour les squelettes ou tests animaux) permet de retrouver, en particulier par le « principe des connections » d'Étienne Geoffroy-Saint-Hilaire (sont homologues des organes ou structures qui, sur deux organismes différents, présentent les mêmes connexions avec les organes ou structures qui les entourent).
Une illustration simple est l'homologie des membres chiridiens antérieurs : nageoire de Panderichthys - membre d'Acanthostega - aile de Ptérodactyle - aile de chauve-souris - aile d'oiseau.

Source - © 1997 La Recherche n°296, p.60
1- Aile de chauve-souris Epomorphus gambiensis ; 2- Bras humain, Homo sapiens ; 3- Bras d'alligator, Alligator mississipiensis ; 4- Aile de puffin cendré, Calonectris diomeda.
L'existence de ces homologies, définies ici dans leur sens non darwinien, s'explique facilement en admettant un ancêtre commun pour les formes qui partagent une même homologies, ancêtre d'autant plus ancien que le nombre d'organismes possédant la même homologie est grand. L'homologie prend alors sa définition darwinienne de structure identique partagée par deux organismes en raison de l'existence d'un ancêtre commun entre eux.
Le fait que les organes homologues ne soient pas pour autant identiques implique une histoire propre pour chaque organisme descendant, depuis l'état de l'ancêtre commun, et amène donc la notion de « descendance avec modification », concept de base de la théorie de l'évolution.
Dernier apport de la paléontologie à la construction de cet édifice théorique : la mise à jour d'organismes qui, sinon constituent, du moins sont probablement très proches de ces « ancêtres communs » hypothétiques.
Paléontologie et formes de transition
On peut rester, pour illustrer ce point, sur l'exemple d'Acanthostega (avec l'article de La Recherche n°296, p.58-61), qui possédait un mélange de traits "primitifs" de poissons (arches branchiales et branchies fonctionnelles), et de traits "dérivés", "évolués", de tétrapodes (chiridium).
Remarque. Il vaut mieux éviter le terme « chaînon manquant » : aucun fossile n'est à coup sûr la forme ancestrale d'une espèce ultérieure ou actuelle, elle présente toujours ou presque (au moins pour ce qui est des organismes pluricellulaires non planctoniques) des caractères qui lui sont propres ("autapomorphies" des phylogénies cladistiques)
On peut achever l'argumentation par les données suivantes, qui réfutent certaines affirmations créationnistes.
Paléontologie, passage des crises, notion et exemple d'exaptation
Les études poussées des épisodes de crises géologiques et biologiques permettent de montrer qu'au sein d'un groupe touché par l'extinction, on peut trouver une ou plusieurs espèces qui surmontent la crise, et qui peuvent être considérées, de par leur morphologie, comme les ancêtres des formes ultérieures , ou de proches parents de ces ancêtres. C'est particulièrement vrai pour les organismes du nanoplancton. Un exemple peut être la figure 178, p.247 du Lethiers, pour les Foraminifères planctoniques à la limite Crétacé/Tertiaire, ou la figure 150 pour les Trilobites de la limite Frasnien/Famennien.
D'après ces données, aucune création de novo n'est à invoquer après le passage d'une crise. Il en va de même avec les « taxons Lazare », des organismes qui paraissent disparaître à la crise, mais que l'on retrouve ensuite au-delà, signe qu'ils ont survécu, sans doute dans des milieux-refuges restreints.
D'autre part, la découverte de nouveaux fossiles montre parfois qu'un caractère que l'on supposait propre à un groupe d'organismes (caractère-diagnostic), et qui paraissait constituer pour lui une « adaptation-clé » (c'est-à-dire qui aurait assuré son succès dans la compétition écologique par occupation d'une niche écologique nouvelle) se rencontre chez des organismes plus anciens, que d'autres caractères excluent du groupe précédent.
Le cas récent que l'on peut mentionner ici est celui des "dinosaures à plumes" : des dinosaures coureurs portant des plumes sur les membres antérieurs et la queue, mais dont la structure du squelette exclut qu'ils aient pu voler : la plume ne peut plus être considérée comme diagnostic d'un oiseau. Et sa structure complexe rend très improbable l'idée qu'elle soit apparue deux fois indépendamment (d'autant que d'autres arguments morphologiques indiquent des liens de parenté entre dinosaures et oiseaux).
Cet exemple a trois intérêts.
- Il apporte une illustration supplémentaire de l'existence de liens de parenté entre des organismes qui, dans la nature actuelle, paraissent très différents, donc de l'Évolution.
- Il permet de concevoir plus facilement comment l'Évolution peut rendre compte de l'existence d'organes très complexes, le cas d'école étant l'œil des Vertébrés ou des Céphalopodes. On conçoit mieux l'existence d'états plus simples pour ces organes si ceux-ci n'avaient pas forcément pour fonction de voir.
- Il constitue un premier exemple de mécanisme évolutif proposé par la paléontologie : une structure remplissant une fonction (=adaptée) chez certains organismes, et utilisée dans une fonction différente par leurs descendants. Ce phénomène de changement de fonction au cours de l'Évolution est appelé « exaptation ».
 Source - © janvier 2000 Pour la Science |

Ce que la paléontologie montre de l'évolution
Il faut d'abord souligner que l'atout principal de la paléontologie est d'examiner l'Évolution dans son déroulement sur des durées géologiques, qui n'ont rien à voir avec les durées des expériences biologiques de spéciation et sélection artificielle.
Outre l'exaptation décrite plus haut, on va présenter ici plusieurs modalités évolutives mises à jour initialement par les paléontologues, faisant toujours l'objet de débats quant à leur importance respective ou leurs causes, et qui demandent désormais à être confrontées aux données ou aux propositions d'explications d'autres disciplines, comme l'écologie et la biologie du développement.
La spéciation et ses rythmes : les radiations adaptatives
La paléontologie ne peut expliquer les spéciations dans leurs mécanismes, justement parce que sa résolution temporelle est trop faible, et les fossiles souvent trop épars. Pourtant, l'abondance de certains groupes fossiles dans les sédiments et au cours du temps montre d'importantes variations de diversité, avec des événements d'extinctions de grande ampleur, attribuables par d'autres données à des événements extrinsèques de modification du milieu, mais aussi (ce qui nous intéresse ici) avec des épisodes de spéciation intensive sur des durées géologiquement brèves. Pour illustrer ce point, on peut prendre la radiation des Mammifères, après la crise Crétacé/Tertiaire (par exemple Lethiers, Fig.71, p.112). Ce phénomène, nommé radiation adaptative, est interprété comme suit : l' apparition de niches écologiques vacantes permet à un groupe de s'y installer et de se spécialiser dans chacune d'entre elles. Cette spécialisation s'effectue par des modifications morphologiques importantes, et qui apparaissent rapidement.
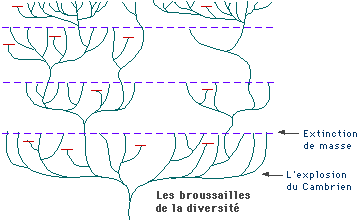
Notons encore que la paléontologie propose deux explications à l'invasion de niches écologiques nouvelles : soit la libération de ces niches par leur occupants précédents, touchés par une extinction, soit l'acquisition, par l'ancêtre du groupe considéré, d'une adaptation-clé, qui lui ouvre la capacité à occuper des niches nouvelles : l'exemple classique ici est celui des Rongeurs, avec leur incisive en rabot à croissance continue.
La paléontologie prouve donc ici que l'évolution s'effectue à des vitesses variables, fonction de l'état du reste du milieu (autres éléments de la biocénose et/ou état du biotope). Elle est aussi à même de montrer les modifications du milieu qui ont eu lieu dans le passé, et de proposer des corrélations entre ces changements et ceux des organismes.
On va voir qu'elle documente aussi l'évolution "de fond", c'est-à-dire la divergence des organismes au cours du temps en dehors des épisodes de grand bouleversement de la biosphère.
Les deux modes d'évolution documentés : anagenèse et équilibres ponctués
Le mécanisme d'anagenèse, le premier conceptualisé et documenté, consiste en une modification graduelle d'une espèce, sur toute son aire de répartition.
En conséquence, la notion d'espèce est malaisée à appliquer ici, puisque le passage d'une forme extrême à l'autre s'effectue par une série continue de transitions. On définit alors les « espèces paléontologiques », ou « chronoespèces », comme suit : connaissant l'étendue moyenne des variations morphologiques actuelles au sein des espèces les plus proches parentes des espèces fossiles considérées, on postulera que l'on passe d'une chronoespèce à la suivante lorsque la variation morphologique entre la forme ancestrale de départ et son descendant sera d'ampleur supérieure à la variation phénotypique intraspécifique constatée sur les formes actuelles.
Les exemples les plus probants de telles évolutions se rencontrent sur les microfossiles planctoniques des séries sédimentaires océaniques. Ces séries sont en effet les plus complètes, et fournissent la meilleure résolution temporelle. Les microfossiles planctoniques, eux, de par leur grande capacité de dispersion, sont les plus à même de maintenir un flux génique sur toute leur aire de répartition, condition indispensable pour une évolution réellement anagénétique. On en trouvera une bonne illustration dans l'ouvrage de J.J.Jaeger, sur le Foraminifère Lepidolina multiseptata, p.104 (ci-dessous).
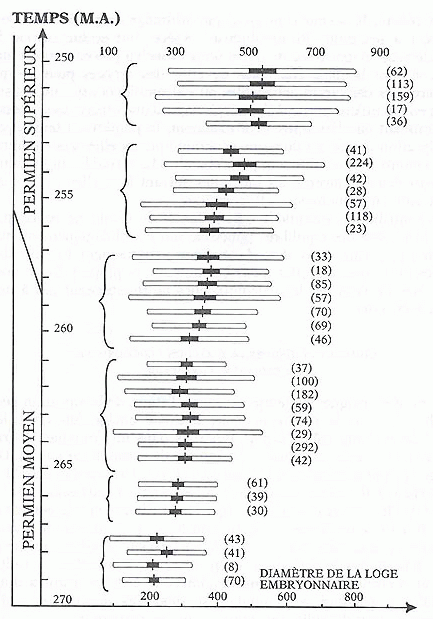
Source - © 1996 J.J. Jaeger, Éditions Odile Jacob
Second mécanisme, d'illustration bien plus récente (Eldredge et Gould, 1977), celui des équilibres ponctués. Ce mode d'évolution suppose que les phénomènes de spéciation s'effectue quasi-instantanément à l'échelle géologique, et sont suivis d'une stase, autrement dit une durée importante pendant laquelle l'espèce ne présente pas de modifications (en tout cas morphologiques, les seules visibles sur le fossile). On trouvera des illustrations de ce mécanisme évolutif dans les livres de Jaeger et de Lethiers (fig.7, p.23).
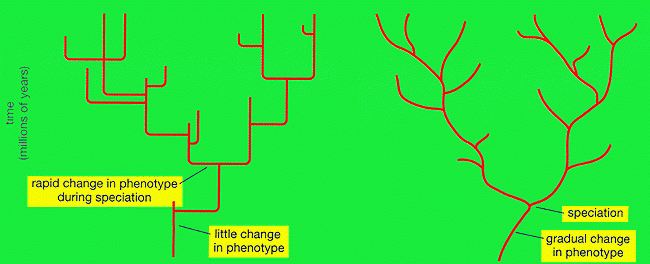
Source - © 1996 J.J. Jaeger, Éditions Odile Jacob
L'existence de cette modalité évolutive est bien établie. Les critiques de cette théorie portent essentiellement sur le mécanisme explicatif proposé par ses inventeurs, à savoir la spéciation allopatrique d'une petite population séparée de la population initiale, qui, après avoir acquis un avantage évolutif, va remplacer la population ancestrale. Les données paléontologiques plus récentes, et les travaux d'écologie sur les espèces actuelles, montrent que les petites populations évoluant en isolats ("îles" au sens large) se révèlent moins compétitives face aux populations de plus grande taille, restées sur le "continent". Aussi le mécanisme explicatif donné plus haut paraît-il improbable.
Ici, la paléontologie ne fait que montrer la modalité évolutive, et proposer des explications. Elle ne peut prouver les mécanismes supposés, et a donc besoin des études d'une autre discipline, ici l'écologie, sur l'actuel. Il en va de même avec l'exemple suivant.
Les hétérochronies du développement
Le principe de ce mécanisme est le suivant : l'espèce descendante présente, à l'état adulte, par rapport à son ancêtre adulte, un ou des caractères morphologiques qui paraissent soit "hyperadultes" (peramorphose), soit caractéristiques d'un état juvénile de l'ancêtre (paedomorphose).
Plusieurs modalités aboutissant à ces résultats sont distinguées (hypermorphose, accélération, pré-déplacement, d'une part, progenèse, néoténie et post-déplacement d'autre part) que l'on ne détaillera pas ici. On peut se contenter d'en illustrer un seul exemple, la néoténie étant la plus documentée. On se référera pour cela aux livres de J.J. Jaeger et de C. Babin, ainsi qu'à l'article de La Recherche de janvier 1999, de J.Chaline et D.Marchand.
Notons enfin que dans de nombreux cas, un organisme présente, par rapport à son ancêtre, des caractères qui paraissent résulter, pour certains, d'un type d'hétérochronie, pour d'autres d'un type différent ("mosaïque" de caractères), ce qui ne facilite pas les tentatives d'explication embryologique.
Là encore, il s'agit d'une modalité d'évolution que la paléontologie illustre, et pour laquelle elle permet d'échafauder des hypothèses de mécanismes explicatifs, sans les prouver. Cette dernière tâche ne peut être remplie que par les biologistes du développement. Cette impuissance de la paléontologie sur certains problèmes constituera l'idée de conclusion.
Conclusion
La paléontologie est par essence une science descriptive, dont on a cependant illustré ici la puissance et la nécessité pour argumenter la théorie de l'Évolution, dans son principe d'abord, dans ses modes d'action ensuite. Il reste qu'elle ne peut ni trouver ni démontrer les mécanismes internes, au niveau génétique et ontogénétique (la traduction du génotype en un ou des phénotypes) de l'Évolution. Elle est par exemple incapable de déceler les phénomènes de spéciation dus à des remaniements chromosomiques, qui introduisent une barrière à la reproduction sans affecter le phénotype (seul élément accessible au paléontologue) (cf. par exemple La Recherche n°296, p.123 et pp.70-75).
Elle ne peut "qu'apporter du grain à moudre" aux biologistes de l'Évolution, et des arguments pour ou contre les hypothèses qu'ils proposent, en particulier en leur fournissant la dimension temporelle que leurs études des organismes actuels ne leur donnent pas. Elle calibre par exemple les hypothèses de divergence entre groupes d'organismes obtenues par les généticiens à partir de l'établissement "d'horloges moléculaires".
En fait, elle fournit une estimation de date pour une séparation donnée entre deux lignées, date à partir de laquelle le généticien établit une horloge moléculaire, qu'il va chercher à utiliser pour d'autres paires de groupes d'êtres vivants, dont la divergence est moins bien documentée par la paléontologie.
De plus, elle illustre la diversité de la vie passée, qui se révèle parfois bien supérieure à ce qui était auparavant imaginé, et le chemin tortueux suivi par la vie pour fournir l'état actuel que nous observons.
Bibliographie et liens
- C. Babin, 1991. Éléments de paléontologie, Armand Colin, 464p. ISBN : 2200210744.
- J.6J. Jaeger, 1996. Les Mondes fossiles, Odile Jacob, 275p. ISBN-13: 978-2738103451.
- F. Lethiers, 1998. Évolution de la biosphère et événements géologiques, Gordon and Breach Science Publishers, 336 . ISBN : 9056991248.
- La Recherche n° 296 Mars 1997 spécial L'Histoire de la vie. 3 milliards d'années d'évolution.
- Examiner aussi les articles récents de paléontologie dans la Recherche ou Pour la Science.
- Dossier Pour la Science n°14, L'Évolution, Janvier 1997.
- M. Ridley, 1997. Évolution biologique, 2e édition, De Bœck Université, 744 p. ISBN : 2804121119.
- Les articles récents des grandes revues anglophones, Nature et Science, pour se tenir au courant.
Les illustrations proviennent de plusieurs sites internet, dont :
- Celui du Muséum de Paléontologie de l'université de Berkeley, en Californie.
- Celui de l'Encyclopaedia Britannica.
- Celui du Muséum National d'Histoire Naturelle.
- Celui de Pour la Science.
Liens utiles sur l'évolution. Ils sont pour la plupart en anglo-américains.
- The TalksOrigins Archives : une mine d'explications sur l'évolution.
- Evolution on the Web for Biology Students : un site de liens par R.Gendron, professeur de biologie à l'Indiana University of Pennsylvania de pour les étudiants en biologie évolutive.
- The Tree of Life web project : un site formidable, qui vous montrera toute la classification des êtres vivants.
- Le site sur l'Évolution de l'Université de Genève : pour ceux qui souhaiteraient maintenant en savoir plus encore sur la théorie de l'évolution du point de vue de la biologie et de la génétique, ou apprendre les bases des hypothèses actuelles sur l'origine de la vie ou de la sexualité.

