
Article | 29/11/2005
Les Poacées (Graminées), nourriture des dinosaures au Crétacé
29/11/2005
Résumé
La découverte en Inde, dans des coprolithes (fèces fossilisées) de Dinosaures, de plusieurs structures siliceuses caractéristiques de différents groupes de Poacées suggèrent (1) que ces plantes étaient déjà présentes et relativement diversifiées dès la fin du Crétacé (alors que les restes fossiles indubitables connus jusqu'alors ne remontaient qu'au Paléocène) et (2) que les Poacées constituaient déjà une part notable de l'alimentation de certains grands dinosaures herbivores.
Table des matières
Une étude publiée dans la revue Science du 18 novembre 2005 [1] vient d'apporter des données inattendues sur deux domaines différents, l'évolution des plantes et l'alimentation des dinosaures herbivores non-aviens. L'analyse de fèces fossiles de grands dinosaures herbivores y a révélé la présence de petites structures siliceuses synthétisées dans les feuilles des Poacées (nom actuel du groupe des Graminées). Cette découverte prouve bien sûr que ces plantes entraient dans le régime alimentaire de ces dinosaures. Mais plus fondamentalement, elle repousse au Crétacé les dates d'apparition et de diversification des Poacées, que l'on ne connaissait jusqu'alors sous forme fossile que depuis le Paléocène. Cette découverte modifie donc les connaissances sur l'histoire évolutive des Poacées et plus largement remet en question les représentations de la végétation du Crétacé.
Les phytolithes des végétaux
Quiconque a déjà arraché machinalement un brin d'herbe ou une tige de graminée sait qu'il est facile de se couper par ce simple geste. Ce caractère abrasif de ces plantes s'explique en partie par le renforcement de leurs tiges ou de leurs feuilles par des dépôts de verre de silice (opale). Cette caractéristique n'est pas l'apanage des graminées ni du groupe des monocotylédones auquel appartiennent les graminées, mais elle y est particulièrement importante. Ce contenu en silice a des conséquences directes sur les mammifères brouteurs, dont les dents sont rapidement usées par cette nourriture abrasive.
La précipitation de silice dans les tissus végétaux
La silice est présente dans tous les végétaux, mais en concentration variable selon les groupes. Elle provient directement de la silice dissoute dans la solution du sol, en général sous forme d'acide silicique (Si(OH4)). Cet acide silicique est pompé par les racines et entre ainsi dans la composition de la sève brute par des mécanismes encore mal connus. Les plantes les moins riches en silice pourraient n'utiliser qu'un mécanisme d'absorption passif (associé à l'absorption d'eau et au taux d'évapo-transpiration). Par contre, certaines plantes plus siliceuses, dont le riz [3] , effectueraient un pompage actif de la silice.
La capture de silice et la quantité de silicium présente dans les différents végétaux sont en effet très variables : les Equisétinées (prêles) sont ainsi très riches en Si, qui imprègnent leur tige. Au sein des Angiospermes, les Monocotylédones sont en général plus riches en silice que les non-monocotylédones [4]. Cette silice précipite sous forme d'opale (SiO2, nH2O), en incrustation dans les parois végétales lignifiées ou sous forme de précipité dans le réticulum endoplasmique, à l'intérieur même de la cellule. Cette imprégnation des parois par l'opale assure donc une fonction de soutien complémentaire de celle de la lignine ainsi qu'une défense contre le broutage. On considère d'ailleurs souvent que cette minéralisation représenterait une adaptation à la pression écologique des herbivores.
Ce renforcement des parois permettrait également une meilleure protection envers les champignons pathogènes [5] et une meilleure tolérance à la dessication. La précipitation de silice permettrait aussi de chélater des métaux toxiques comme l'aluminium ou le zinc [7]. Quoiqu'elle ne soit généralement pas prise en considération dans les compositions des engrais, la silice serait donc pourtant un constituant utile sinon nécessaire à la croissance des plantes.
Les mécanismes de précipitation de la silice sont encore mal connus. Une hypothèse classique considère cette précipitation comme un phénomène passif liée à l'évapo-transpiration, dans la mesure où ces dépôts concernent en effet les régions de la plante où l'évapo-transpiration est la plus forte. Toutefois, Piperno et al. [6] ont constaté sur des cultures de courge (Cucurbita) que le dépôt d'opale ne s'effectuait que dans les cellules à paroi lignifiée. Ces auteurs proposent que, chez cette plante là au moins, la précipitation d'opale soit génétiquement contrôlée par l'intervention d'un locus dominant également impliqué dans le dépôt de lignine. Cependant, l'acide silicique présente une grande affinité pour les composés poly-hydroxylés qui entrent en jeu dans la synthèse de la lignine, ce qui pourrait aussi expliquer cette corrélation entre lignification et silicification [7].
Les dépôts de silice, en moulant les tissus qu'ils imprègnent, forment des structures appelées phytolithes (ou phytolites, "pierre de plante"), dont la forme varie selon celle de la cellule et donc, finalement, selon la plante qui les produit. Ils peuvent ainsi devenir des éléments de diagnostic taxonomique et prouver la présence d'un groupe de plante donné dans l'environnement. Les phytolites sont par conséquent devenus un objet d'étude pour les archéologues, qui tentent grâce à eux de reconstituer les environnements mais aussi les régimes alimentaires des populations passées.
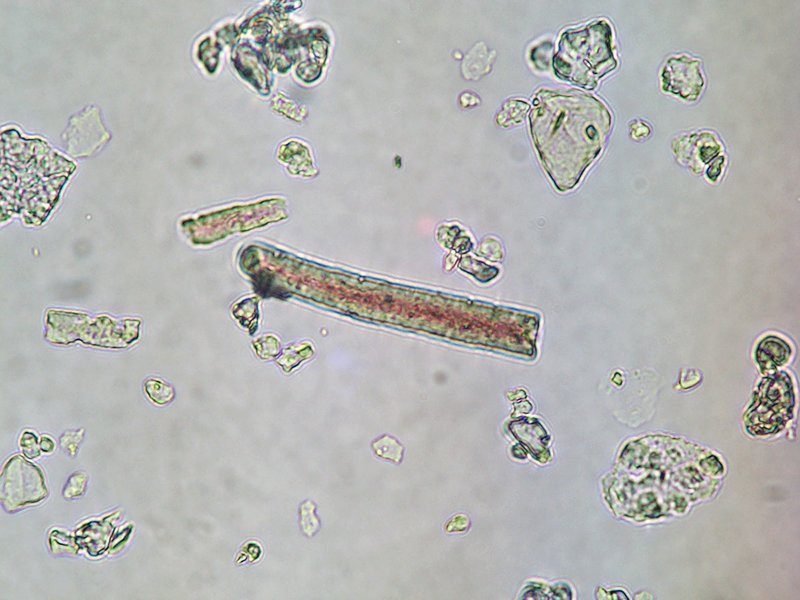
Source - © 2005 H.-G. Naton
Figure 1. Phytolithes allongés vus au microscope
Les phytolithes des Poacées, outils de détermination phylogénétique
L'examen des phytolithes des Poacées actuelles a montré que certaines morphologies de phytolithes sont spécifiques de grands sous-groupes monophylétiques de Poacées. Ces phytolithes peuvent donc diagnostiquer la présence de ces Poacées dans les environnements, selon la même démarche que celle suivie par la palynologie : certains morphotypes s'avèrent caractéristiques de l'ensemble des Poacées à l'exception des lignées les plus basales, d'autres d'un premier grand sous-clade regroupant les bambous, le riz (Oryza), l'orge, l'avoine (Avena) et le blé (Triticum) -- clade BEP (Bambusoidés/Ehrhartoidés/Pooidés) -- d'autres encore du second grand sous-clade des Poacées (clade PACCAD), comprenant entre autres les actuels Mais (Zea) et le roseau (Phragmites).
 Source - © 2005 Cyril Langlois Figure 2. Bambous en croissance, Nord du Vietnam | 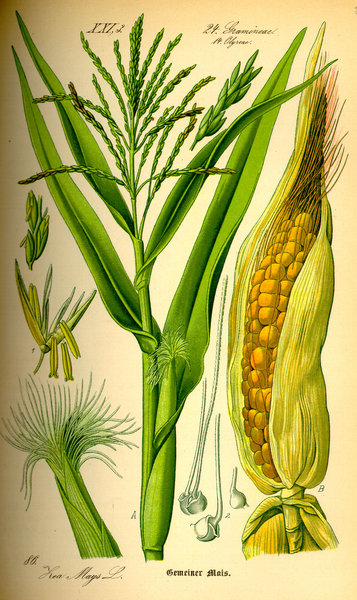 Source - © 1885 O.W. Thomé, Flora von Deutscland, Österreich und der Schweiz Figure 3. Zea maïs, le maïs actuel |
Le bambou et le maïs, deux représentants respectifs actuels (parmi beaucoup d'autres) des clades BEP et PACCAD.
Les découvertes de Prasad et al.
Prasad et al. [1] ont analysé des coprolithes fossiles (c'est-à-dire des fèces fossilisées) provenant de Pisdura, au centre de l'Inde, un site situé en bordure des trapps du Deccan, ces immenses plateaux de laves d'âge voisin de la limite Crétacé-Tertiaire. Le site étudié serait donc daté du Crétacé très récent (Maestrichtien, vers -67 Ma). La présence dans les mêmes niveaux de fossiles de grands dinosaures herbivores non-aviens (Titanosauridés) conduit ces auteurs à attribuer ces fèces à ces animaux.
Les Titanosauridés étaient des dinosaures sauropodes herbivores et quadrupèdes (proches morphologiquement et phylogénétiquement des fameux Brachiosaures et Diplodocus du Jurassique). Ils se distinguaient de ces derniers par un revêtement dorsal de plaques osseuses. Ce clade était apparemment présent sur tous les continents à la fin du Crétacé et ce groupe a sans doute compté les plus grands vertébrés terrestres qui aient jamais existé, certaines espèces approchant peut-être les 100 tonnes [9].
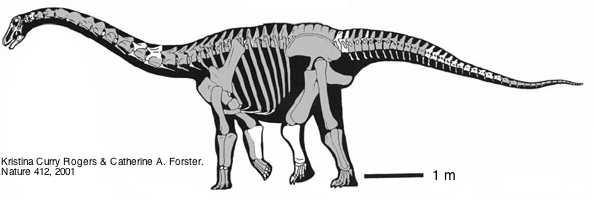
Source - © 2001 K. Curry Rogers et C.A. Forster, Nature
Seuls les éléments squelettiques figurés en blancs ont été effectivement retrouvés. La barre d'échelle représente un mètre.
Les auteurs ont dissout les coprolithes pour en extraire les microfossiles siliceux, phytolithes et diatomées d'eau douce en particulier. L'examen de ces restes leur a permis de caractériser plusieurs morphotypes de phytolithes et de les rapprocher des morphotypes connus chez les plantes actuelles. Ils ont ainsi découvert des phytolithes provenant probablement pour certains de Conifères, pour d'autres de palmiers, et surtout plusieurs morphotypes attribuables à des Poacées. Certains morphotypes ou certaines associations de morphotypes sont rapprochés des phytolithes des clades de Poacées actuels. La figure ci-après montre la correspondance obtenue entre certains morphotypes, indiqués par des carrés noirs et des flèches à droite, et une phylogenèse des Poacées dérivée de la phylogénie des Poacées actuelles proposée par le Groupe de Travail sur la Phylogénie des Herbacées (Grass Phylogeny Working Group, un consortium de chercheurs auteur en 2001 d'une phylogénie des Poacées actuelles).
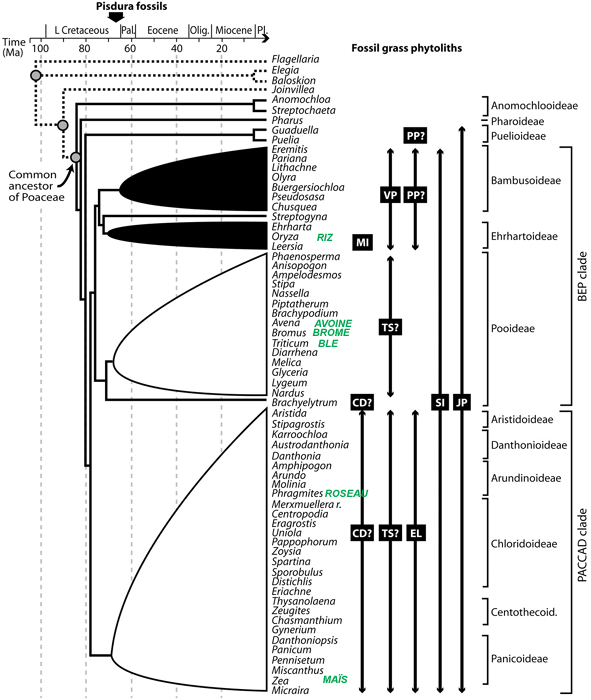
Source - © 2005 V. Prasad et al., Science
Voir le texte pour plus d'information. Sont ajoutées en vert les noms de quelques Poacées actuelles courantes.
Jusqu'à présent, les macrofossiles de Poacées indubitables ne remontaient qu'au Paléocène (~ 55 Ma). Seuls des pollens attribués à des herbes de ce groupe permettaient de supposer qu'il existait des Poacées depuis la fin du Crétacé sur les continents de l'hémisphère Sud qui formaient l'ancien continent du Gondwana. À l'inverse, les études phylogénétiques faisant appel au principe de l'horloge moléculaire proposaient des dates plus anciennes (~ 83 Ma) pour l'apparition de ces Angiospermes. Les découvertes de ces auteurs confirment donc cette existence des Poacées à la fin du Crétacé. Mais de plus, les phytolithes retrouvés montrent déjà une large diversité et, de plus, aucun d'eux n'est attribuable aux groupes les plus "basaux" des Poacées, ce qui signifie que la diversité du Groupe étaient déjà élevée au Maestrichtien et que les deux sous-clades majeurs comptaient plusieurs représentants à cette époque.
Quelques implications de ces nouvelles données
Des Poacées dans le régime des dinosaures... et de qui d'autres ?
Les phytolithes récupérés dans ces déjections fossiles peuvent y avoir été incorporées par ingestion inconsciente de débris de plantes ou de terre mêlés à la nourriture, ou par une consommation volontaire des plantes productrices. Selon Prasad et al., étant donné que les morceaux d'opale récupérés présentent des états de préservation variables, les deux types d'ingestion devaient être à l'œuvre et ces Sauropodes pouvaient prélever activement des Poacées. Néanmoins, la nourriture consommée était variée : conifères, cycadales, dicotylédones et herbes monocotylédones. Étant donné que les Monocotylédones actuelles contiennent beaucoup plus de silice que les Dicotylédones et les Conifères, le nombre relativement modeste de phytolithes de Monocotylédones récupérés implique que ces herbes ne constituaient pas une part majeure de l'alimentation de ces dinosaures.
D'ailleurs, les dinosaures herbivores du Crétacé ne présentent aucune adaptation dentaire à la consommation de végétaux abrasifs et riches en silice. À l'inverse, nombre de mammifères brouteurs d'herbes du Tertiaire disposent de telles adaptations, en particulier l'hypsodontie : augmentation de hauteur de la couronne dentaire, qui prolonge la durée d'utilisation de la dent malgré son usure progressive. Cependant cette adaptation des Mammifères résulte aussi de ce que ce groupe ne dispose que de deux générations de dents (dents de lait et dents définitives). Chez les dinosaures non-aviens, comme chez les crocodiles, des dents étaient formées régulièrement et les dents usées, brisées ou perdues étaient rapidement remplacées, l'hypsodontie ne leur aurait donc pas apporté d'avantage évolutif particulier, surtout si les herbes ne représentaient qu'une partie de leur régime alimentaire.
Par contre, il existait à la fin du Crétacé et au Paléocène un petit groupe de Mammifères encore assez mal connu, les Gondwanathères, rapprochés ou inclus dans le groupe éteint des Multituberculés. Ces mammifères présentent des dents typiquement hypsodontes et des représentants de ce groupe figurent parmi les fossiles trouvés en Inde dans des formations équivalentes à celles étudiées par Prasad et al. Il n'est donc pas exclu que ces animaux aient été des brouteurs d'herbes dès la fin du Crétacé et qu'une coévolution entre herbacées dures et herbivores brouteurs ait déjà été à l'œuvre, même si cette hypothèse reste très spéculative.
Les paysages du Crétacé : quelle place pour les Poacées ?
Une autre implication des données de Prasad et al. concerne la reconstitution et la représentation des paysages du Crétacé récent. Jusqu'à présent, les représentations montraient des dinosaures évoluant dans des paysages dominés par les Conifères et les Cycadales. Il pourrait s'avérer nécessaire de réviser ces illustrations si la part réelle des Poacées dans les paysages étaient effectivement plus importante que ce que l'on supposait jusque là et si des pelouses à Poacées existaient déjà dans certaines parties du Gondwana.
L'histoire évolutive des Angiospermes : une chronologie et une géographie à réviser ?
L'histoire évolutive des Angiospermes est encore peu connue. Que les deux grands sous-clades de Poacées soient déjà relativement diversifiés au Crétacé récent tend à réconcilier les données fossiles et moléculaires sur une apparition de ce groupe au cours du Crétacé, peut-être vers -86 Ma. La diversification des Poacés se serait alors produite rapidement durant le Crétacé récent et une vingtaine de millions d'années aurait suffi à l'apparition de représentants des deux sous-clades.
L'autre aspect de l'évolution des Poacées auquel cette étude apporte des éléments supplémentaire concerne l'origine géographique du groupe : les cartes ci-après rappellent la géographie de la fin du Crétacé.
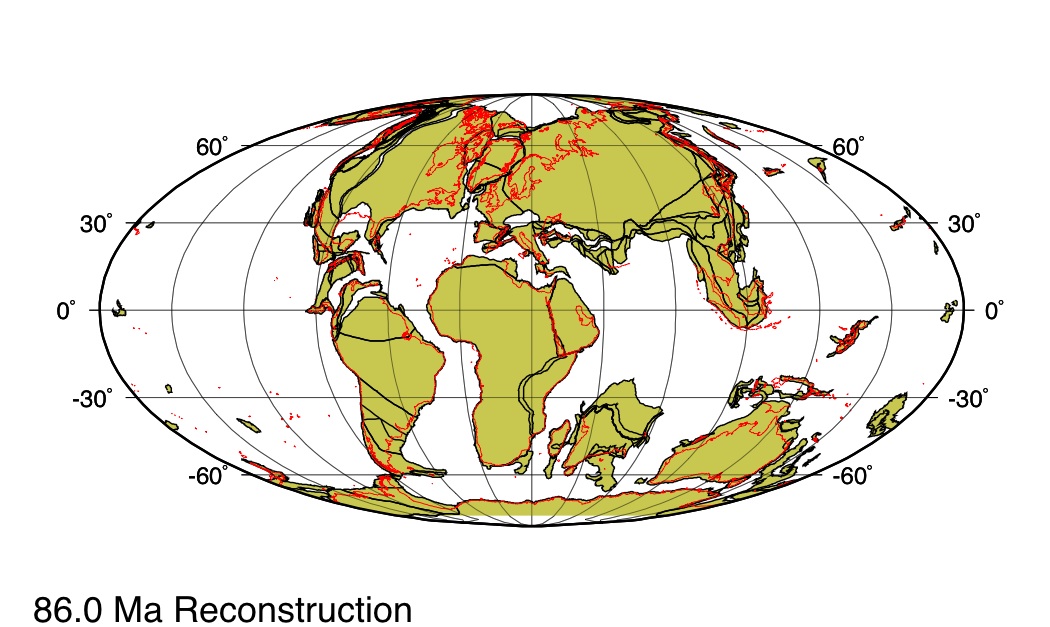 Source - © 2005 ODSN Figure 6. Répartition des continents il y a 86 Ma Les contours actuels des terres émergées sont indiqués en rouge. | 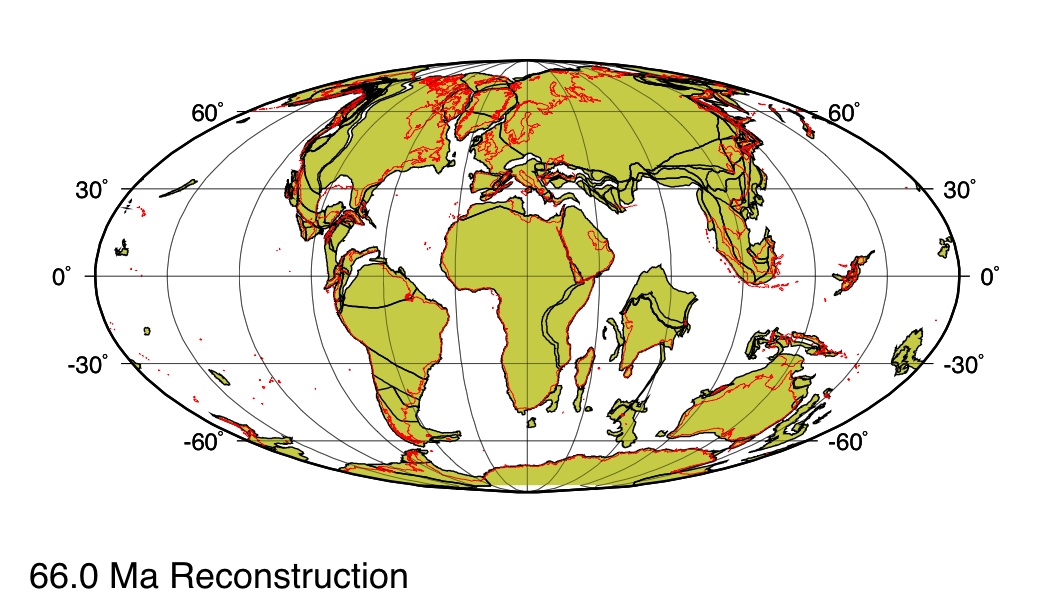 Source - © 2005 ODSN Figure 7. Répartition des continents il y a 66 Ma Les contours actuels des terres émergées sont indiqués en rouge. |
- Une première hypothèse proposait que les Poacées soient apparues en Asie avant de s'étendre sur l'Inde puis sur le reste des continents Sud qui représentaient l'ancien Gondwana. Cette hypothèse est considérablement affaiblie puisque la disposition des continents au Crétacé récent exclut tout contact entre l'Inde, alors isolée, et l'Asie (figures ci-dessus).
- Une deuxième possibilité serait l'apparition du groupe en Inde puis son expansion sur les continents de l'hémisphère Nord, Asie puis Amérique du Nord, au cours du tertiaire. Selon Prasad et al., cette hypothèse n'est que peu soutenue, car les pollens d'herbes du Paléocène - Éocène sont beaucoup plus rares dans le registre fossile des continents septentrionaux que dans celui des continents méridionaux.
- Dernière solution, favorisée par les auteurs, une origine "gondwanienne" des Poacées, sur l'un des continents de l'hémisphère Sud, suivi d'une diversification du groupe soit par dispersion des populations sur des masses continentales relativement séparées soit par vicariance (diversification par division de la population initiale du fait de la séparation des masses continentales). Les Poacées auraient pu, dans ce cas, apparaître en Inde ou ailleurs sur les terres du Gondwana avant la complète dislocation de ce supercontinent. le point important est que les deux sous-groupes (BEP et PACCAD) devaient déjà avoir une extension géographique suffisante pour se trouver en Inde lorsque ce bloc a définitivement perdu ses connexions avec les autres masses gondwaniennes.
Conclusion
Cette étude de débris siliceux microscopiques récupérés dans des fossiles d'ordinaire bien peu "médiatiques" amènent donc des résultats importants et soulèvent de nouvelles questions. Certaines, comme le régime alimentaire des Gondwanathères ou la place des Poacées dans les paysages du Crétacé, ne sont pour l'heure que des conjectures ou des pistes de recherche à approfondir. Le résultat principal reste l'extension dans le Crétacé du registre fossile des Poacées et le rapprochement qu'il permet avec les résultats de phylogénie moléculaire. Il souligne encore l'utilité fondamentale de la recherche d'échantillons sur le terrain, seule à même d'apporter des données concrètes en faveur ou en défaveur des hypothèses formulées à partir d'autres approches (phylogénie, géochimie, etc). Ces travaux ont aussi l'intérêt de rappeler que les paléontologues tentent de "faire parler" tous les matériaux fossiles à leur disposition et par tous les moyens disponibles. Si des moyens nouveaux comme la géochimie des isotopes stables sont de plus en plus mis à profit, ils ne font que s'ajouter à une panoplie d'approches plus "classiques" mais indispensables.
Bibliographie
- Prasad V., Strömberg A. E. et al. , Dinosaur coprolites and the evolution of grasses and grazers. Science vol. 310, 18 novembre 2005
- Piperno, Dolores R. et Hans-Dieter Sues, Dinosaurs dined on grass. Science vol. 310, 18 novembre 2005
- Mitani M. et Ma, J., Uptake system of silicon in different plant species. Journal of Experimental Botany 56 (414), 2005
- Hodson, M. J. et al., Phylogenetic variation in the silicon composition of plants. Annals of Botany, 2005
- Hammerschmidt, R. Silicon and plant defense: the evidence continues to mount. Physiological and Molecular Plant Pathology 66(5), 2005
- Piperno Dolores R. et al., Evidence for the control of phytolith formation in Cucurbita fruits by the hard rind (Hr) genetic locus: Archaeological and ecological implications. PNAS 99(16), 2002
- Richmond Kathryn E. et Sussman Michael, Got silicon? The non-essential beneficial plant nutrient. Current Opinion in Plant Biology 6(3), 2003
- ODSN. Make plate tectonic reconstruction. http://www.odsn.de/odsn/services/paleomap/paleomap.html
- Benton M. J., Vertebrate Palaeontology, 2nd edition. Blackwell, 2000.

