Image de la semaine | 15/01/2018
Welwitschia mirabilis, une Gnétale unique du désert de Namibie
15/01/2018
Résumé
De Welwitschia mirabilis, espèce endémique de Namibie et d'Angola, au clade des Gnétales, discussion sur les classifications morphologique et phylogénétique des Spermatophytes et caractère paraphylétique des Gymnospermes.

Source - © 2001 Pierre Thomas
Figure 1. Welwitschia mirabilis au milieu du désert namibien
La personne située à côté montre la taille imposante de la plante.
 Source - © 2001 Pierre Thomas Figure 2. Welwitschia mirabilis au milieu du désert namibien Vue de plus près, on voit une importante masse constituée par les feuilles, ainsi que des cônes reproducteurs (strobiles) mâles qui dépassent. |  Source - © 2001 Pierre Thomas Figure 3. Welwitschia mirabilis au milieu du désert namibien Vue de plus près, on voit une importante masse constituée par les feuilles, ainsi que des cônes reproducteurs (strobiles) femelles qui dépassent. |
Welwitschia mirabilis est réputée pour être l'une des plantes endémiques les plus étranges au monde. Ses pieds poussent au milieu du désert côtier de Namibie et d'Angola, ainsi que dans des secteurs désertiques arbustifs plus à l'intérieur des terres (non illustrés dans le présent article), où ils forment des amoncellements verts pouvant atteindre de très grandes tailles.
Lorsqu'on les observe de près, on constate que ces amoncellements sont en fait constitués de seulement deux feuilles en ruban produites par un méristème basal. Ces feuilles sont insérées sur un tronc très court et souterrain. Elles ont une croissance continue et peuvent pousser jusqu'à atteindre des très grandes tailles (entre 2 et 4 m). La croissance est lente. On estime que certains plants ont plus de 1000 ans. Au cours de cette croissance, ces deux feuilles sont dilacérées en lanières plus étroites, ce qui conduit à l'aspect final des plantes qui semblent avoir plusieurs feuilles étroites et longues.
 Source - © 2001 Pierre Thomas Figure 4. Détail d'un "jeune" plant de Welwitschia mirabilis On voit sur ce cliché la base des deux feuilles rubanées. Elles forment des rubans entiers à la base et qui sont dilacérés par l'extrémité terminale au cours de la croissance. Certains font l'hypothèse que la forme en entonnoir de la partie aérienne de la plante permettrait de concentrer l'eau de la rosée vers la racine pivotante. |  Source - © 2001 Pierre Thomas Figure 5. Détail d'un très jeune plant de Welwitschia mirabilis Sur un plant très jeune, on voit bien les deux feuilles, qui sont encore peu dilacérées. |
 Source - © 2001 Pierre Thomas Figure 6. Détail d'un plant de Welwitschia mirabilis Sur ce plant plus âgé, on devine encore la symétrie bilatérale due aux deux uniques feuilles, et on voit que les tiges reproductrices partent de la base des feuilles. |  Source - © 2017 Pierre Thomas Figure 7. Détail de la zone de croissance d'un plant de Welwitschia mirabilis Du fait de la présence de deux seules feuilles, la zone de croissance a typiquement une symétrie bilatérale en forme d'amande. Derrière, on distingue du bois de gymnosperme permien silicifié (cf. Une "forêt pétrifiée" de gymnospermes permiens du Damaraland, près de Khorixas, Namibie). |
 Source - © 2017 Pierre Thomas Figure 8. Détail de la zone de croissance d'un plant de Welwitschia mirabilis Sur certains plants jeunes, la symétrie n'est pas visible et le méristème basal a une forme plutôt circulaire (mais un fonctionnement symétrique). Des morceaux de bois fossilisé permien ont été disposés pour éviter son piétinement. | |
Les organes reproducteurs de Welwitschia mirabilis sont organisés en strobiles (des cônes regroupant les structures reproductrices) situés sur des tiges fertiles. L'espèce est dioïque, un plant donné produisant soit des cônes mâles, soit des cônes femelles.
 Source - © 2017 Pierre Thomas |  Source - © 2001 Pierre Thomas Figure 10. Détail d'un pied mâle de Welwitschia mirabilis avec des strobiles mâles On reconnaît les strobiles mâles par leur côté foisonnant et, au moment de la floraison, par leurs étamines. |
 Source - © 2011 KLEIN Benjamin , CC BY-SA-4.0 Figure 11. Détail d'un cône mâle de Welwitschia mirabilis en floraison On distingue bien des étamines de couleur rouge. Le cône est formé de plusieurs structures mâles agglomérées. La pollinisation est assurée par des insectes. | |
 Source - © 2017 Pierre Thomas |  Source - © 2001 Pierre Thomas |
 Source - © 2001 Pierre Thomas | |
Welwitschia mirabilis est la seule espèce du genre Welwitschia. Ce genre appartient à un clade, les Gnétales, dont les trois seuls genres actuels sont Welwitschia, Gnetum et Ephedra.
Les Ephedra constituent un genre d'espèces xérophytes (adaptées aux conditions de sécheresse). Elles ont un port de genêt, avec une hauteur variable selon les espèces et les conditions de croissance (de quelques centimètres à quelques décimètres de haut). En France, on en trouve sur la côte Atlantique avec Ephedra distachya (aussi appelée raisin de mer). Cette espèce est présente du Portugal à l'Asie centrale, toujours en conditions de sécheresse relative. Ephedra gerardiana est une espèce qui pousse dans l'Himalaya. Les Ephedra font partie de la pharmacopée traditionnelle indienne. On en extrait aujourd'hui l'éphédrine, un vasoconstricteur qui a un effet similaire à l'adrénaline.
 Source - © 2016 Marc Coudel Les graines sont contenues dans une enveloppe charnue rouge, qui est comestible (au moins à faible dose). Comme chez Welwitschia, les structures reproductrices forment des strobiles (c'est à dire qu'elles sont concentrées sur une portion terminale de rameau où elles forment un cône). | |
 Source - © 2011 Marc Coudel Figure 16. Touffe d'Ephedra gerardiana dans l'Himalaya proche de l'Everest (4500 m d'altitude environ) C'est l'une des dernières plantes ligneuses observée à cette altitude dans le massif. Auteur: Marc Coudel 2011 |  Source - © 2011 Marc Coudel Figure 17. Vue rapprochée d'Ephedra gerardiana Elle ressemble à un Equisetum, mais ne présente pas de ramifications en verticilles. Marc Coudel 2011 |
Les Gnetum sont des lianes qui poussent en milieu tropical.

Source - © 2005 BotBln, CC BY-SA-3.0
Figure 18. Gnetum gnemon, une liane d'Asie du Sud-Est
Les Gnétales possèdent un certain nombre de caractères dérivés (caractères "nouveaux" issus de la modification de caractères ancestraux au cours de l'évolution aboutissant au taxon considéré). Parmi eux, certains sont particulièrement intéressants car les Angiospermes présentent des caractères dérivés avec des ressemblances importantes :
- Le bois. Le bois est hétéroxylé (il possède des vaisseaux vrais, et pas seulement des trachéides comme les Pinophytes) et sa lignine donne une couleur rouge intense après actions successives du permanganate de potassium et de l'ammoniac.
- La fécondation est double. Deux spermatozoïdes produits par le grain de pollen sont conduits par le tube pollinique et fusionnent avec des noyaux du gamétophyte femelle. Contrairement aux Angiospermes, un seul des zygotes formés se développe tandis que l'autre dégénère (chez les Angiospermes, le deuxième zygote donne le périsperme, un tissu nourricier pour l'embryon).
L'ovule est entouré d'une enveloppe charnue. Cela ressemble à première vue à un fruit charnu.
Rappelons que le fruit est une structure propre aux Angiospermes. Sa partie charnue dérive alors d'un ou plusieurs carpelles. Elle contient une ou plusieurs graines, qui dérivent d'un ou plusieurs ovules. Chaque graine est enveloppée par deux téguments (comme chez les Conifères). Un carpelle est parfaitement clos. Par conséquent, au moment de la fécondation, les grains de pollen ne peuvent être déposés directement au niveau de l'ovule et la croissance du tube pollinique commence dès le stigmate.
Chez les Gnétales, l'enveloppe charnue n'est séparée du reste de la graine que par un seul tégument. De plus, elle est discontinue. Elle laisse en effet passer un micropyle allongé. Cela permet le dépôt des grains de pollen directement au contact de l'ovule où le tube pollinique peut croître. Ces arguments montrent que d'un point de vue structural, l'enveloppe charnue des Gnétales est le tégument externe de la graine !

Source - © 2007 Marco Schmidt, CC BY-SA-2.5
Figure 19. Feuilles et ovules de Gnetum africanum
Chez les Gnetum, les feuilles sont plates avec des nervures ramifiées et ressemblent beaucoup à celles de certaines Angiospermes. Il y a une enveloppe charnue (rouge chez cette espèce) autour de la graine. On distingue sur le cliché la sortie du micropyle, ce qui différencie ces structures des fruits clos des Angiospermes.
Source - © 2017 Marc Coudel
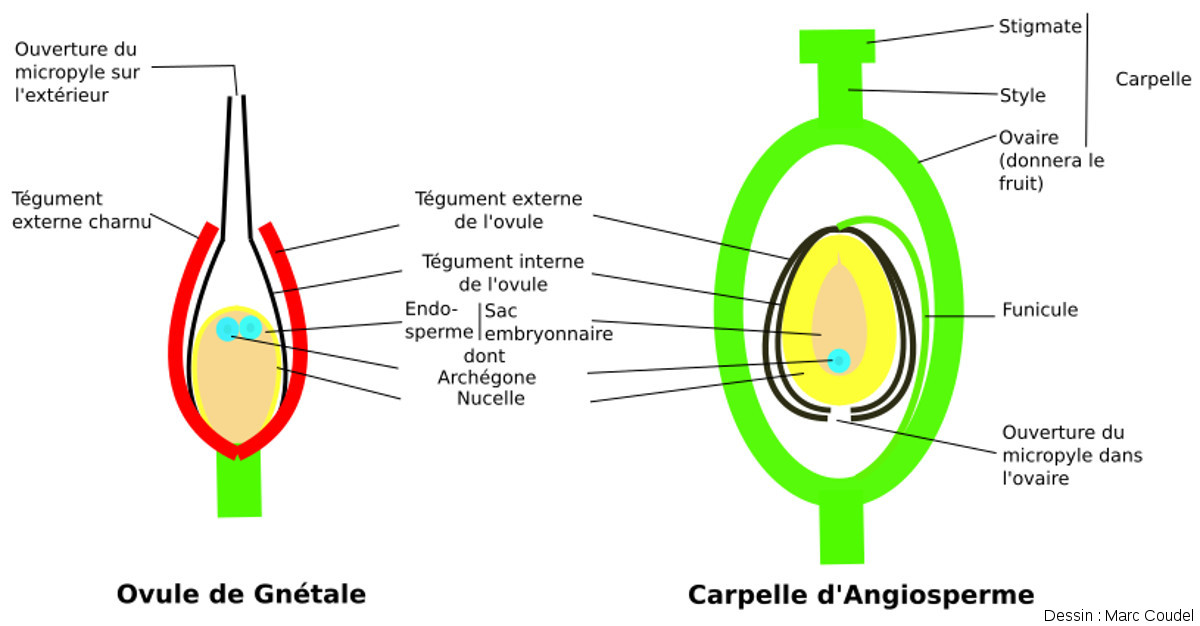
Figure 20. Schéma comparatif entre les ovules de Gnétales et le gynécée d'une Angiosperme
L'ovule est la structure qui, suite à la fécondation, devient une graine. Il a une structure similaire chez les deux groupes. Chez les Gnétales, c'est l'un des téguments de l'ovule qui est charnu. Le micropyle donne directement sur l'extérieur, donc l'enveloppe est discontinue et on ne parle pas de fruit. Chez les Angiospermes, l'ovule est lui-même contenu dans un ovaire clos à l'intérieur duquel s'ouvre le micropyle. Après la fécondation, la paroi de l'ovaire se transforme en fruit, qui est complètement fermé. Il y a alors angiospermie totale (la graine est dans une “urne”).
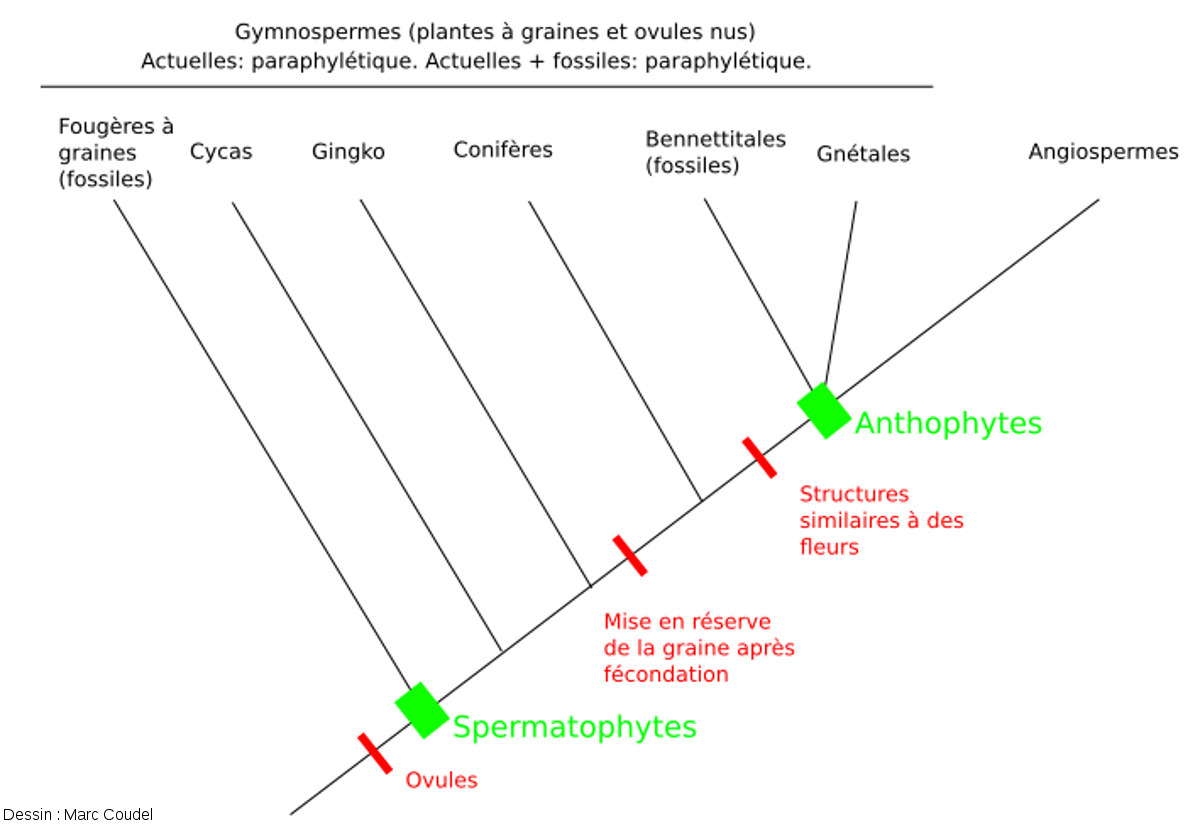
Les premières classifications des Gnétales s'appuyaient sur les caractères morphologiques. Comme les caractères évoqués dans la partie précédente ressemblaient beaucoup à ceux des Angiospermes, on a supposé leur homologie pour former le groupe des Anthophytes. Ce groupe comprenait aussi les Bennettitales, des plantes fossiles (cf. Feuilles de Ptéridospermales et de Bennettitales du Kimméridgien, Cerin (Ain)). Selon cette hypothèse, les Gymnospermes actuelles (plantes à graines et ovules nus comprenant les Cycadales, les Gingkoales, des Conifères et les Gnétales) formaient un groupe paraphylétique.
Source - © 2017 Marc Coudel
Selon cette hypothèse, les Gnétales formeraient avec les Angiospermes le groupe monophylétique des Anthophytes. Les groupes formés par les Gymnospermes actuelles et par les Gymnospermes actuelles + fossiles sont paraphylétiques.
Outre les erreurs de positionnement des diverses bifurcations, erreurs parfaitement compréhensibles avant les études moléculaire, cette représentation telle qu’on la faisait dans les décennies précédentes est maladroite, voire tendancieuse. Il y a sur ce schéma une ligne droite qui va des premiers Spermaphytes jusqu’aux Angiospermes. Tous les autres groupes s’écartent de cette ligne droite et semblent n'en être que des rameaux latéraux "imparfaits". Sans que ce soit explicitement dit ni même voulu, cela laisse implicitement penser (à un élève par exemple) que les Angiospermes ont une place à part chez les végétaux, que ce sont les plus évolués et accomplis des végétaux, voire qu’ils découlent d’un dessein, d’un dessein intelligent (intelligent design).
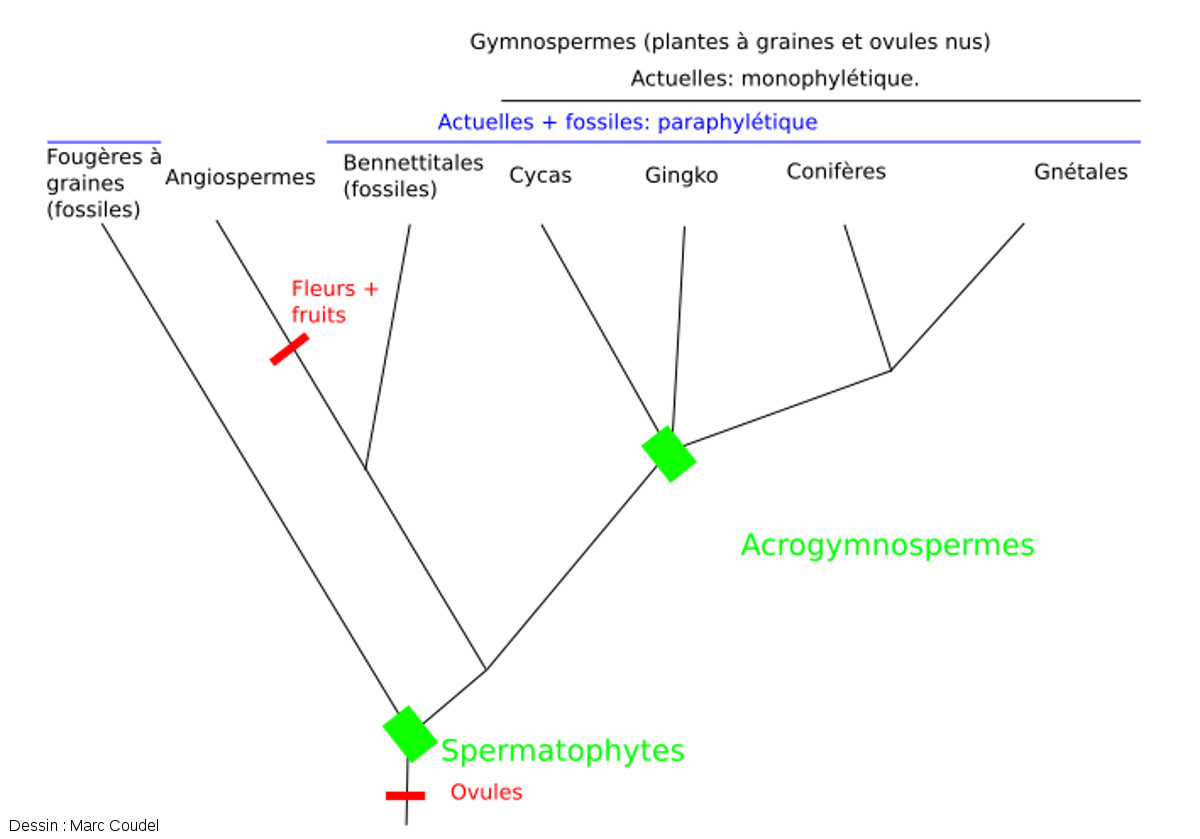
Cependant, les classifications moléculaires récentes ont complètement bouleversé les classifications des plantes. Les nouvelles données montrent que les ancêtres des Angiospermes ont divergé des ancêtres des Gymnospermes actuelles de façon précoce. C'est probablement aussi le cas des Bennettitales, fossiles pour lesquels on ne dispose pas de données moléculaires. Les Gymnospermes actuelles forment donc un groupe qui est monophylétique ! On l'a appelé Acrogymnospermes. Si on inclut les taxons fossiles (et donc les Bennettitales), les Gymnospermes restent cependant un groupe paraphylétique.
Ces classifications sont très robustes. On peut en conclure que les nombreux caractères morphologiques similaires entre les Gnétales et les Angiospermes sont analogues plutôt qu'homologues ! Cela illustre la difficulté inhérente à l'utilisation des caractères morphologiques dans les classifications de la Lignée verte du fait des nombreuses convergences.
Source - © 2017 Marc Coudel
Figure 22. Classification actualisée des Spermatophytes basée sur des phylogénies moléculaires
Dans cette hypothèse, les Gymnospermes actuelles forment un groupe monophylétique. Il s'agit des Acrogymnospermes. Il regroupe les Conifères, ainsi que les Cycadales et Gingkoales (auparavant considérées comme des divergences basales) et les Gnétales (auparavant considérées comme Anthophytes). En revanche, si on ajoute des clades fossiles (Bennettitales), les Gymnospermes forment toujours un groupe paraphylétique.
Aujourd'hui, la relation entre les Gnétales et les Conifères est encore discutée. Il est en effet possible que les Gnétales forment un groupe ayant divergé au sein de celui des Conifères plutôt qu'un groupe qui en aurait divergé précocement.
Toutes les photographies de P. Thomas de cet article ont été prises lors d'une excursion géologique organisée par le CBGA (Centre briançonnais de géologie alpine) et encadrée par Olivier Dauteuil (Université de Rennes).