Article | 09/06/2006
Tiktaalik, un nouveau témoin de la transition poissons Sarcoptérygiens - Tétrapodes
09/06/2006
Résumé
Un nouveau fossile de poisson Sarcoptérygien découvert dans l'arctique canadien vient se placer en parent immédiat des premiers Tétrapodes du Dévonien. Il complète ainsi l'histoire que l'on pouvait dresser jusqu'alors des étapes évolutives qui ont conduit à l'acquisition des caractéristiques anatomiques des Tétrapodes. L'excellente conservation des spécimens retrouvés permet de reconstituer ce proche cousin des Tétrapodes et de montrer que ses caractéristiques anatomiques, quoiqu'intermédiaires entre celles des "poissons" et celles des "tétrapodes terrestres", n'en faisait pas moins un animal fonctionnel et efficace eu égard à son mode de vie. Sur le plan pédagogique, ce fossile permet donc d'aborder les problèmes conceptuels liés aux notions de "chaînon manquant" et d'acquisition de caractères nouveaux.
Table des matières
Cet article a été relu et validé par Lionel Cavin du département de géologie et de paléontologie du Muséum d'histoire naturelle de Genève.
Des poissons Sarcoptérygiens aux Tétrapodes
Les Sarcoptérygiens du Dévonien
La parenté des Tétrapodes et des poissons Sarcoptérygiens est depuis longtemps proposée et a été étayée par la découverte de Sarcoptérygiens et de Tétrapodes fossiles très proches par certaines caractéristiques anatomiques, les os des membres en particulier. Un poisson Sarcoptérygien fossile en particulier, Eusthenopteron, d'âge Dévonien supérieur, sert de base de comparaison pour les fossiles de tétrapodes ou de "proto-tétrapodes". Chez cet animal, les nageoires sont charnues et constituées de plusieurs os formant un axe. L'attache avec les ceintures scapulaires et pelviennes se fait par un seul os, homologue de l'humérus et du fémur. On reconnaît déjà, dans la patte antérieure, les os homologues de ceux du membre chiridien des tétrapodes, à l'exception des doigts.
Les premiers Tétrapodes
On dispose aujourd'hui de plusieurs fossiles de Tétrapodes primitifs plus ou moins complets. Deux d'entre eux sont particulièrement bien connus aujourd'hui et illustrent déjà la relative diversité de ce groupe à la fin du Dévonien : Acanthostega et Ichthyostega. Comme dans le cas du nouveau fossile décrit ici, aucun de ces animaux ne représente un véritable "chaînon manquant". Ce sont des animaux particuliers, possédant un mode de vie propre et des caractéristiques anatomiques spécifiques, mais dont certains caractères anatomiques peuvent, a posteriori, être vus comme des "transitions" entre les états de leurs homologues plus anciens et plus récents.
Ces Tétrapodes archaïques possédaient tous les deux :
- un membre chiridien complet, mais avec plus de cinq doigts ;
- un crâne large, aplati dorso-ventralement, aux os soudés, donc plus résistant mais aussi plus léger que celui des poissons sarcoptérygiens, et séparé de la ceinture scapulaire : l'animal a un cou, des muscles étaient fixés à l'arrière du crâne et permettaient à la tête, devenu d'un seul tenant et allégée, de se relever et de s'abaisser, y compris hors de l'eau où la poussée d'Archimède est négligeable ;
- des côtes orientées latéralement et élargies, abritant probablement des poumons comme chez les actuels Dipneustes ; mais ces animaux disposaient toujours de branchies, comme leurs parents les poissons sarcoptérygiens ; par contre, puisque la tête n'était plus fixée à l'épaule, ces branchies n'étaient plus recouvertes par un opercule osseux ;
- un museau allongé et une mâchoire inférieure aux os eux aussi soudés, un trait nécessaire à une ventilation pulmonaire efficace. Il permet en effet d'aspirer l'air et de l'envoyer aux poumons par dilatation et contraction de la cavité buccale, un mécanisme toujours utilisé par les Amphibiens et les Dipneustes actuels.
- Le stapes (appelé étrier dans l'anatomie humaine), os de l'oreille moyenne qui dérive de l'os hyomandibulaire situé, lui, dans la cavité branchiale des poissons Sarcoptérygiens. La présence de cet étrier ne signifie cependant pas qu'il assurait déjà l'audition en milieu aérien comme il le fait chez les Tétrapodes terrestres plus récents.
- Un "système de ligne latérale", organe sensible aux variations de pression dans l'eau, formant une ligne sur le flanc de l'animal et un réseau de canaux sensoriels sur le crâne.
- Une queue allongée, élargie par des expansions des vertèbres et bordée d'une nageoire soutenue par des rayons dermiques homologues des rayons des poissons.

Source - © 1997 J.A Clack, La Recherche,296
Ces animaux étaient donc à même de respirer hors de l'eau, mais tous les deux devaient avoir néanmoins un mode de vie essentiellement aquatique.
- Dans le cas d'Acanthostega, cette conclusion est d'autant plus probable que l'orientation des membres de cet animal ne lui permettait pas de soutenir son corps ou de le soulever pour compenser la gravité en milieu aérien.
- Ichthyostega, lui, avait des membres antérieurs assez puissants et musclés et une différenciation des vertèbres qui suggèrent que les muscles qui s'y attachaient étaient spécialisés pour différents mouvements. On peut imaginer que cet animal se déplaçait sur le rivage en prenant appui sur ses pattes avant et en tirant l'arrière de son corps par une ondulation verticale. Mais plusieurs autres caractères, comme par exemple les pattes rappelant plus des "palmes", démontrent qu'Ichthyostega restait un animal beaucoup plus efficace en milieu aquatique [J. A. Clack, PLS, février 2006].
Tiktaalik rosae, le nouveau venu
Le site fossilifère et le matériel retrouvé
L'expédition à l'origine de la découverte de Tiktaalik avait pour objectif précis la recherche de sarcoptérygiens susceptibles de documenter les étapes évolutives survenues entre les Tétrapodes primitifs déjà connus et les Sarcoptérygiens plus anciens. L'équipe a donc recherché des terrains fossilifères du Dévonien moyen et supérieur. Le site de fouille finalement sélectionné se trouve aujourd'hui au sud de l'île d'Ellesmere, dans le territoire canadien du Nunavut (figure 2).
Les spécimens retrouvés ont été extraits de grès fins datés du Dévonien supérieur (Frasnien), correspondant à des sédiments déposés dans les méandres peu profonds d'un grand delta fluviatile. Ce delta se situait, au Dévonien moyen et supérieur, sur la bordure nord de la Laurasia, un continent alors situé sur l'équateur. Plusieurs autres fossiles de Sarcoptérygiens apparentés aux Tétrapodes proviennent de régions situées à l'époque sur le pourtour de ce même continent (figure 3).
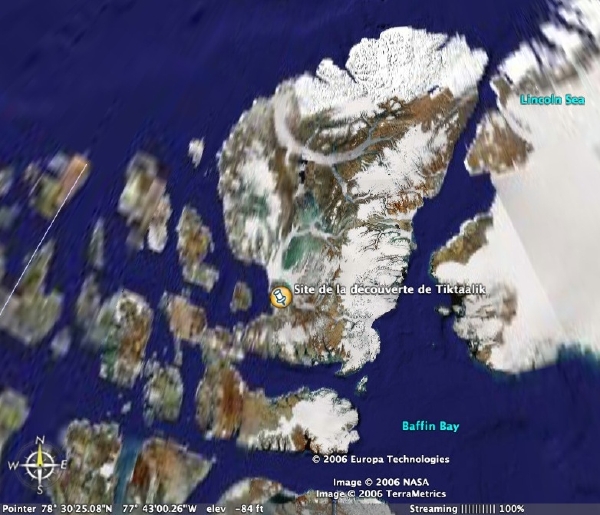 Source - © 2006 Google earth | 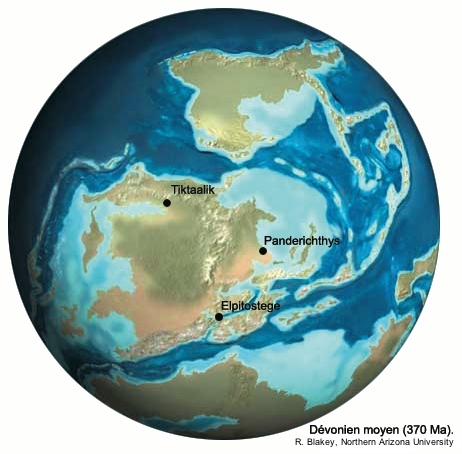 Source - © 2006 R. Blakey, Northern Arizona Univ. La carte représente la géographie du Dévonien moyen (370 Ma), elle est donc légèrement antérieure aux fossiles considérés, mais la géographie d'ensemble reste valable pour le Dévonien supérieur. |
Le site fossilifère d'Ellesmere a livré de nombreux éléments du squelette de Tiktaalik, par exemple plusieurs échantillons de mandibule, correspondant à au moins dix individus, et des animaux plus complets, comprenant encore le crâne et la colonne vertébrale en connexion, remarquablement bien préservés. Ces individus, conservés en trois dimensions et seulement légèrement écrasés, se trouvent "dispersés dans toute la région fossilifère, se recouvrant souvent et sans orientation préférentielle" [Daeschler et al. (2006)]. On dispose ainsi d'une image assez précise de cet animal.
Les caractéristiques de l'animal
L'animal découvert présente plusieurs traits semblables à ceux des Tétrapodes dévonien déjà connus et plus récents.
- Le crâne est large et aplati dorso-ventralement.
- Les yeux sont situés sur le sommet du crâne.
- Le museau est long comparativement à la longueur totale du crâne.
- Les côtes sont élargies et orientées latéralement.
Dans le même temps, il reste un "poisson", avec de petites nageoires pelviennes, des nageoires portant des rayons, des branchies et, au moins sur la peau du dos, partiellement préservée, un revêtement d'écailles.
Mais Tiktaalik prend également une place de "transition" dans la phylogénie des Sarcoptérygiens par l'anatomie de son membre antérieur, que ses découvreurs ont pu décrire en détail [Shubin et al., (2006)]. La reconstitution de ce membre suggère que cette patte pouvait non seulement adopter l'orientation classique des nageoires des Sarcoptérygiens non-tétrapodes connus jusqu'ici, mais qu'elle pouvait également se plier et se tourner partiellement vers l'avant au niveau du "coude". Le radius et la fibula (le cubitus) avaient également une relative indépendance : "l'avant-bras" pouvait donc également tourner légèrement par rapport au "bras". Cette mobilité permettait à l'animal de poser ses nageoires antérieures sur un substrat, dans une position para-sagittale, et donc de s'appuyer sur elles (figure ci-dessous). Cela n'implique pas directement la capacité à marcher ni une vie en milieu aérien : seuls les membres antérieurs de Tiktaalik rosae présentent cette capacité à reposer sur le sol, les membres postérieurs sont des nageoires charnues plus courtes, orientées plus ou moins horizontalement ; si l'animal pouvait se déplacer sur un support solide, et peut-être hors de l'eau, à l'aide de ces nageoires — ce que suggèrent ses découvreurs — c'était à la façon d'un phoque.

Source - © 2006 D'après Shubin et al., Nature, modifié
Figure 4. Reconstitution des positions possibles de la nageoire antérieure de Tiktaalik rosae
A : étendue horizontalement pour la nage ; B : pliée et appuyée sur le substrat.
Hormis ce membre antérieur, la morphologie de Tiktaalik présente d'autres traits particuliers, comme la forme très aplatie du museau, et la position légèrement surélevée des narines et des orbites, qui rappelle l'aspect des crocodiles. Cette forme et le membre antérieur conduisent ses découvreurs à reconstituer cet animal stabilisé sur le fond d'un cours d'eau peu profond par ses nageoires, les yeux et les narines au ras de l'eau, à l'instar des crocodiliens à l'affût.

Source - © 2006 Ted Daeschler, Univ. Chicago
Figure 5. L'un des exemplaires les plus complet du nouveau Sarcoptérygien tétrapodomorphe Tiktaalik rosae
Cette vue de trois-quart avant souligne la longueur du museau, la forme générale aplatie du crâne et du corps (sur le spécimen présenté, le crâne est probablement peu déformé, donc naturellement aplati, même si le corps, lui, a été écrasé).
Les fossiles retrouvés ne permettant pas de reconstituer la partie la plus postérieure de l'animal, on ignore s'il disposait d'une large nageoire caudale, comme Acantosthega, et donc s'il pouvait se propulser efficacement et rapidement dans l'eau. On remarquera aussi que si Tiktaalik pouvait se soulever sur ses nageoires antérieures, il était alors "plus adapté" à la locomotion à terre qu'Acanthostega. Sous cet aspect, donc, Acanthostega, quoique plus "tétrapode" par ses membres pourvus de doigts et par d'autres caractères, était peut-être plus nageur que ce nouveau fossile et paraît ainsi plus "archaïque" sur le plan de la locomotion. Le "degré d'évolution" anatomique ne reflète donc pas une "progression" vers l'adaptation au milieu terrestre.
Tiktaalik dans la phylogénie des Sarcoptérygiens
Les caractères morphologiques de Tiktaalik rosae énumérés précédemment permettent de le placer, dans une phylogénie des Sarcoptérygiens dévoniens, en groupe-frère immédiat des Tétrapodes, donc évolutivement plus proche de ces derniers que le fossile Panderichthys, qui représentait jusqu'alors l'un des "plus proches poissons parents" des Tétrapodes. L'absence de doigts ne fait pas de Tiktaalik un Tétrapode, mais il en a le museau allongé et a perdu la couverture osseuses des branchies, contrairement à Panderichthys. Plutôt que comme Tétrapode à part entière, on le décrira plutôt comme un "Sarcoptérygien tétrapodomorphe". Sur la phylogénie obtenue à partir de ces fossiles de poissons et d'amphibiens dévoniens, Tiktaalik prend la même place qu'un autre fossile, Elpitostege, déjà décrit auparavant mais représenté seulement par quelques éléments de mâchoire.

Source - © 2006 P.E. Ahlberg, J.A. Clack, 2006, Nature
Figure 6. Position de Tiktaalik dans la phylogénie des Sarcoptérygiens tétrapodomorphes et des Tétrapodes
La figure représente cinq des animaux les mieux connus : l'ostéolépiforme Eusthenopteron, les deux formes "transitionnelles" Panderichtys et Tiktaalik et les deux Tétrapodes primitifs Acanthostega et Ichthyostega. Notez sur les schémas des toits crâniens, à gauche, i) la position dorsale des yeux chez Tiktaalik et les Tétrapodes qui suivent ii) la disparition des os recouvrant les branchies (en bleu), déjà absents chez Tiktaalik, et iii) la réduction de longueur des os post-pariétaux (en vert) ainsi que l'allongement relatif du museau.
Conclusion : des "étapes" évolutives résultats d'adaptations spécifiques diverses
Comme on l'a vu, Tiktaalik rosae est une découverte importante par son état de conservation exceptionnel et parce qu'il prend effectivement un statut de "fossile de transition" ou de "chaînon" au moins aussi important qu'Archaeopteryx. Ce faisant, il constitue un exemple intéressant pour expliquer les notions d'état de caractère dérivé (apomorphie) et primitif (plésiomorphie) et la méthode cladistique de reconstruction phylogénétique.
Dans le même temps, il illustre aussi les problèmes posés par la prise en compte des fossiles quand il s'agit d'adapter les classifications systématiques aux phylogénies : parmi les fossiles présentés ici, lesquels placer dans les Tétrapodes ? Lesquels considérer comme groupe-frère ? On peut donc insister là auprès des élèves sur la nécessité qu'il y a à considérer, de préférence, des groupes monophylétiques, seuls groupes naturels, mais de la "facilité" qu'il y a à considérer des groupes paraphylétiques comme "les poissons sarcoptérygiens", groupe qui exclurait les Tétrapodes.
Finalement, Tiktaalik est un exemple parmi d'autres pour insister sur la relative artificialité des concepts de "chaînon manquant" et "d'étapes évolutives" dans l'acquisition de certains caractères adaptatifs et, évidemment, sur l'absence totale de toute "finalité" dans ce processus. Ces "étapes" n'apparaissent qu'a posteriori, en comparant des organismes phylogénétiquement proches, mais d'âges différents et souvent d'habitat et de mode de vie eux aussi différents.
Néanmoins, ce fossile montre tout de même que l'on connaît bien certaines transitions importantes dans l'histoire des vertébrés grâce aux fossiles (de la même façon que pour la transition dinosaures - oiseaux). Cette notion contredit l'idée que les fossiles ne permettent pas d'illustrer les "grandes étapes" de l'évolution.
- Dans certains cas, on peut obtenir une série d'organismes qui présentent une adaptation de plus en plus spécialisée à un mode de vie. Ainsi, les plus anciens fossiles attribués au groupe des Cétacés, comme Pakicetus, qui étaient encore des animaux terrestres (avec peut-être un mode de vie proche du raton-laveur ou de la loutre), ont des narines situées au bout du museau. Les Cétacés pélagiques actuels ont les narines au sommet du crâne, ce qui représente une adaptation mammalienne à la vie en milieu strictement aquatique. Or, on connaît effectivement des Cétacés fossiles pélagiques chez qui les narines sont positionnées au milieu du museau, "à mi-chemin" des deux dispositions "ancestrale" et "évoluée". Dans ce cas précis, on pourrait évoquer une spécialisation au cours du temps.
- Avec ces fossiles de Sarcoptérygiens tétrapodomorphes et de Tétrapodes primitifs, on peut voir l'acquisition d'un membre antérieur articulé — permettant à l'animal d'appuyer sa patte ou sa nageoire sur un substrat pour se stabiliser — comme une adaptation de ce groupe d'animaux carnivores à la vie dans des eaux marécageuses peu profondes, encombrées de végétation et éventuellement mal oxygénées.
- Cependant, si l'on considère l'acquisition des caractères de Tétrapodes (le membre marcheur, les osselets de l'oreille moyenne, les doigts, etc.), cette série de fossiles ne montrent pas forcément une "spécialisation progressive" à la vie en milieu émergé, mais plutôt une série de caractères acquis plus ou moins séparément par des animaux différents, en adaptation à des modes de vie particuliers. C'est un cas typique d'évolution "en mosaïque".Les doigts d'Acanthostega n'étaient pas une adaptation à la marche, mais ont été mis à profit ensuite pour ce faire. Ces caractères ont été hérités par leurs ancêtres et réutilisés, re-sélectionnés en tant qu'adaptation à d'autres contraintes. Ce sont des exaptations au sens du paléontologue Stephen Jay Gould, de la même façon que l'utilisation des plumes pour le vol chez les Oiseaux est une exaptation par rapport aux dinosauriens non-aviens chez qui les plumes servaient peut-être de protection thermique.
On remarquera enfin — par exemple avec la disposition horizontale des membres d'Acanthostega, ou la locomotion à terre d'Ichtyostega — que disposer de caractères de Tétrapodes amphibiens — les doigts, le membre chiridien — ne signifie pas pour autant un mode de vie ou une locomotion identiques à ceux des amphibiens actuels — Acanthostega nageait, Ichtyostega traînait peut-être occasionnellement son corps sur le sol avec un mouvement qui évoquerait plus celui d'une chenille que celui d'une salamandre [J. A. Clack (2006)]. Ressemblance anatomique et ressemblance de mode de vie sont deux choses distinctes.
Annexe : Tiktaalik et Archaeopteryx sont-ils équivalents comme "fossiles de transition" ?
La position phylogénétique particulière de Tiktaalik rosae, qui vient compléter l'histoire de l'apparition des premiers Tétrapodes, n'est pas sans évoquer le cas d'Archaeopteryx lithographica, souvent considéré comme un fossile de transition entre les dinosauriens non-aviens et les Oiseaux.
Comme Archaeopteryx lithographica, ce nouveau fossile présente une mosaïque de caractères à l'état primitif (écailles, ligne latérale, nageoires rayonnées, branchies, etc) et de caractères à l'état dérivé (anatomie et articulation du membre antérieur, position des yeux et des narines, proportions du crâne, etc). La comparaison est donc justifiée.
Toutefois, du point de vue de la classification systématique, comme on l'a mentionné précédemment, Tiktaalik n'est pas considéré comme un Tétrapode au sens strict, mais comme un Sarcoptérygien tétrapodomorphe : il ne possède pas encore l'ensemble des caractères utilisés pour définir un Tétrapode et il se place donc en groupe-frère de ceux-ci. Archaeopteryx, par contre, n'est plus considéré comme un dinosaure proche des oiseaux, mais bien comme un oiseau au sens strict. En effet, les analyses minutieuses effectuées sur les fossiles disponibles ont montré que :
- Les plumes des ailes et de la queue d'Archaeopteryx avaient un profil dissymétrique, ce qui représente une adaptation aérodynamique, impliquant leur implication dans le vol. Les plumes des ailes étaient des rémiges, longues plumes recouvrantes formant une surface portante.
- L'anatomie d'Archaeopteryx suggère que cet animal pouvait avoir des muscles suffisamment puissants pour effectuer un vol battu, actif, même s'il ne disposait pas du sternum élargi et de la fourchette flexible qui améliore l'efficacité du vol battu chez les oiseaux plus récents.
- Des analyses récentes, par tomographie, de l'endocrâne d'un des spécimens d'Archaeopteryx, suggèrent (1) que cet animal disposait d'un cerveau relativement volumineux eu égard à sa taille, comme les oiseaux actuels, et (2) qu'il possédait les spécialisations sensorielles (vision performante, sens de l'équilibre, intégration sensorimotrice) impliquées, chez les oiseaux actuels, dans le contrôle du vol [Dominguez Alonso et al., (2004)].
Ces résultats conduisent à considérer Archaeopteryx comme un oiseau à part entière. Dit autrement, le groupe monophylétique des Oiseaux (Aves) est défini comme l'ensemble des animaux à plumes capables d'un vol battu actif (au moins primitivement) et leur ancêtre commun, ce qui inclut donc Archaeopteryx dans cet ensemble.
Certes, cette distinction entre Tiktaalik et Archaeopteryx reste assez mineure et découle des définitions, relativement arbitraires, des groupes monophylétiques des Tétrapodes et des Oiseaux. Comme on l'a dit précédemment, ces deux fossiles restent des exemples frappants de la capacité des fossiles à éclairer le déroulement de certaines étapes évolutives majeures.
Références
- Ahlberg E. and Clack J. A., 2006, A firm step from water to land, Nature 440, 747-749.
- Clack J. A., 1997, Les premiers tétrapodes vivaient dans l'eau, La Recherche n°296, mars 1997.
- Clack J. A., 2006, Le premier pied à terre, Pour la Science
- Dominguez Alonso P. et al., 2004, The avian nature of the brain and inner ear of Archaeopteryx, Nature 430, 666-669. (En ligne sur le site de l'auteur)
- Daeschler E. B. et al., 2006, A Devonian tetrapod-like fish and the evolution of tetrapod body plan, Nature 440, 757 - 763.
- Clément G., 2006, Les tétrapodes de paléo-Belgique, Pour la Science
- Shubin N. et al., 2006, The pectoral fin of Tiktaalik rosae and the origin of the tetrapod limb, Nature 440, 764 - 771.
- Tiktaalik rosae : Site de l'université de Chicago consacré à Tiktaalik rosae et à l'expédition à l'origine de la découverte.