
Article | 02/10/2008
Quand la paléontologie revoit ses données
02/10/2008
Résumé
Analyses de grandes bases de données de fossiles à l'aide d'outils mathématiques, statistiques et informatiques récents. Deux articles de l'été 2008 conduisent à réviser des conclusions et des idées plus anciennes : ils rappellent ainsi combien les progrès des sciences historiques comme la paléontologie ou la paléoclimatologie reposent sur l'accumulation des fossiles mais aussi sur l'amélioration des critères de classification et des méthodes d'analyse de ces données.
Table des matières
Cet article a été aimablement relu et amendé par Loïc Villier, maître de conférence dans le groupe Géologie des Sytèmes et des Réservoirs Carbonatés de l'Université de Provence (Aix-Marseille 1), et membre du projet international "The Paleobiology Database".
Introduction
L'un des principaux thèmes de la paléontologie est l'étude de l'histoire de la biodiversité au cours des temps géologiques, au moins au cours des derniers 540 Ma qui constituent le Phanérozoïque. Ce type d'étude se base, fondamentalement, sur l'analyse des catalogues de fossiles répertoriés dans les collections et, désormais, sur les banques de données qui en prennent le relais.
Mais ces banques de données s'enrichissent au fil du temps, à mesure des découvertes. Elles sont également modifiées ou corrigées au gré des nouvelles déterminations de taxa, des redéfinitions de taxa et des analyses phylogénétiques. Enfin, les techniques d'analyse de ces grands corpus de données, fondées sur des méthodes statistiques rigoureuses et sur l'exploitation de la puissance de calcul des ordinateurs, ont elles aussi rapidement évoluées depuis une cinquantaine d'années.
Deux articles parus pendant l'été 2008, portant sur des sujets a priori assez différents — la biodiversité des Dinosauriens au cours du Crétacé et la biodiversité marine sur l'ensemble du Phanérozoïque — illustrent tout deux comment l'amélioration des bases de données et l'utilisation de techniques d'analyse statistique modernes permettent de re-vérifier de précédents résultats et de remettre en question certaines conclusions plus anciennes.
Ces deux exemples illustrent aussi la modernité de la paléontologie, qui emploie aujourd'hui les méthodes de recherche, les techniques et les technologies les plus récentes : travail d'équipe bénéficiant de la mise en commun des résultats de nombreux chercheurs, bases de données volumineuses et rendues accessibles par Internet, analyses statistiques et phylogénétiques devenues possibles grâce à la puissance des calculateurs modernes.
Les "super-arbres" et les radiations des dinosauriens
La première de ces études, par Graeme T. Lloyd et co-auteurs, a consisté à réaliser un arbre phylogénétique très complet, portant sur les espèces (et non sur les genres ou sur une autre catégorie taxinomique de plus haut rang), du clade des Dinosauriens dans son ensemble (Oiseaux exceptés). En confrontant cet arbre de relations entre taxa au registre fossile de ces mêmes taxa, ils ont pu proposer des dates pour les principaux nœuds de la phylogénie, ce qui leur a ensuite permis d'analyser les variations de diversité du groupe au cours de son histoire.
La technique des "super-arbres" phylogénétiques
Les Dinosauriens — au sens classique, non cladistique, n'incluant pas les Oiseaux — constitue l'un des groupes de vertébrés éteints les mieux connus et les plus documentés. Ce taxon comprend plusieurs grands sous-clades, certains très diversifiés : les grands Sauropodes quadrupèdes (Diplodocus, Brachiosaurus...), les Cératopsiens (quadrupèdes à collerette osseuse cornues, comme Triceratops), les Théropodes (bipèdes carnivores, avec les fameux Tyrannosaurus, Deinonychus ou Velociraptor), etc. L'obtention d'un arbre phylogénétique consensuel pour l'ensemble des dinosaures (même en laissant de côté les Oiseaux), impliquerait l'élaboration d'une matrice taxon-caractères monstrueuse, un nombre d'arbres possibles gigantesque et un temps de calcul inacceptable.
Suivant une démarche déjà utilisée pour obtenir des phylogénies à l'échelle de très grands groupes (la phylogénie des Eucaryotes ou les relations entre Bactéries, Archæbacteries et Eucaryotes, par exemple), cette équipe a donc utilisé la technique des "super-arbres" (supertrees), qui consiste, fondamentalement, à combiner en un seul arbre plusieurs phylogénies obtenues indépendamment pour les différents sous-clades. Cette technique n'est toutefois pas aussi simple que pourrait le suggérer la phrase précédente.
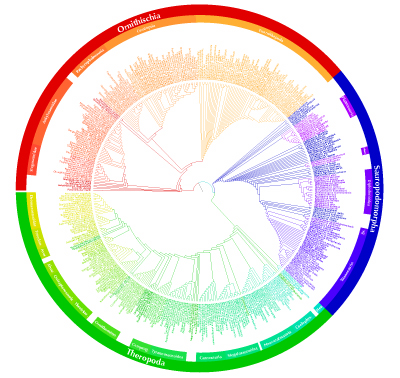
Source - © 2008 Univ. of Bristol, communiqué de presse
Une version PDF haute résolution de ce super-arbre des Dinosauriens (et de figures complémentaires) est disponible sur le site de l'équipe.
En premier lieu, il faut homogénéiser la systématique des espèces en identifiant correctement les espèces valides et leurs éventuels synonymes et en excluant les taxons douteux ou improprement décrits. Il s'agit donc de mettre à jour la nomenclature. Chaque espèce valide a pu être attribuée à l'un des différents genres.
Ensuite, la méthode des super-arbres ne consiste pas à rabouter plusieurs arbres correspondant chacun à un clade entier et complet, mais à recalculer un arbre synthétique à partir d'arbres plus petits (arbres-sources) qui se recouvrent partiellement les uns les autres, autrement dit qui partagent certains de leurs taxa. Les chercheurs ont donc dû reprendre chaque arbre-source pour les rendre compatibles entre eux et adaptés au calcul d'un super-arbre. Chacun de ces arbres est alors transformé en une matrice "taxon-position par rapport à un nœud" : chaque nœud interne significatif de l'arbre est référencé et devient une colonne de la matrice ; tous les taxa situés "en aval" de ce nœud sont affectés de l'état "1", tous les autres reçoivent l'état "0" dans cette colonne.
Toutes les matrices ainsi obtenues sont alors concaténées en une seule matrice à partir de laquelle le super-arbre peut être calculé et optimisé (Bininda-Edmonds, 2004). Ce qui a signifié, ici, le traitement de 155 arbres-sources précédemment publiés et 5000 heures de calcul pour le super-ordinateur de l'Université Nationale d'Irlande, à Maynooth.
L'analyse de l'évolution d'un clade à partir de sa phylogénie
Le super-arbre obtenu a ensuite été converti en une phylogénie, c'est-à-dire replacé dans le temps à l'aide du registre fossile connu des taxa. Cette phylogénie permet d'estimer les variations de diversité du groupe des Dinosauriens au cours du temps. Les épisodes remarquables de diversification ou d'extinction seront facilement identifiables en comparant la forme de l'arbre phylogénétique à celle d'un arbre où la vitesse de renouvellement des faunes resterait constante.
Ce raisonnement relativement simple est, en pratique, insuffisant. Pour avoir une idée plus correcte des changements de diversité du groupe, il faut, par exemple, tenter de corriger les biais d'enregistrement liés aux biais d'échantillonnage. En effet, certaines périodes sont mieux représentées que d'autres sur le terrain, ou les fossiles y sont mieux préservés, ou encore les efforts de fouilles y ont été plus intenses, ce qui peut amener à des surestimations de la biodiversité par rapport à d'autres périodes. Une manière de procéder est de tenir compte des "lignées fantômes" et des "taxons fantômes" (ghost lineages et ghost taxa,Norell, 1992) dans l'estimation de la biodiversité. À partir des phylogénies, on constate que les occurrences, dans le registre fossile, de deux groupes-frères A et B, peuvent être très différentes. Si l'un des deux groupe-frères (A, par exemple) apparaît bien plus tôt que l'autre dans les roches, cela pourrait impliquer que le deuxième, B :
- soit existait déjà avant sa première apparition dans les sédiments ("lignée fantôme") ;
- soit était alors représenté par des organismes qui ne possédaient pas encore les apomorphies diagnostiques de ce taxon. Son groupe, son taxon, existait donc déjà ("taxon fantôme").
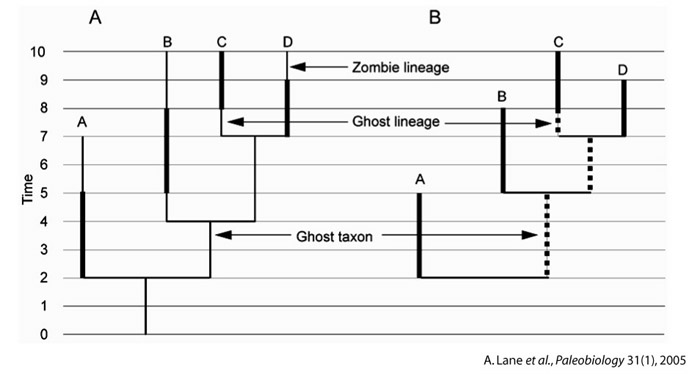
Source - © 2005 Lane et al., Paleobiology 31 (1), modifié
Figure 2. Méthode de correction des extensions temporelles réelles des taxa à partir de leur phylogénie
En A, une phylogénie de quatre taxa et leurs lignages ancestraux, avec leurs extensions temporelles complètes représentées en traits fins. Les traits épais représentent les parties réellement échantillonnées de ces extensions, de la première à la dernière occurrence du taxon dans le registre fossile. Des exemples de "lignées fantômes", avant la première apparition, de "lignées zombies", après la dernière apparition, et d'un "taxon fantôme", non échantillonné, sont représentés. En B, les relations entre taxa sont utilisées pour retrouver certaines portions des extensions temporelles fantômes, indiquées en tirets épais. À l'inverse, les "lignées zombies" des taxa terminaux A, B et D, ne peuvent pas être décelés. Les "taxa fantômes" (ghost taxon) sont créés pour représenter le lignage ancestral des groupes couronnes (le clade (B-C-D), ici).
Dans un cas comme dans l'autre, cela peut signifier que des organismes, "reliant" A et B, existaient en même temps que A mais avant que B soit présent dans le registre fossile : il convient donc de les comptabiliser dans la biodiversité, même s'ils ne sont pas documentés à l'état fossile.
Là encore, ce raisonnement est à appliquer avec précaution et peut même introduire des erreurs de décompte : si le groupe A est, en fait, réellement l'ancêtre de B, et non son groupe-frère (en particulier dans le cas d'une spéciation par "équilibre ponctué", où l'espèce ancestrale A persiste inchangée après qu'une nouvelle espèce B s'en soit détachée), mais que cette relation ancêtre-descendant n'est pas décelée, l'existence du taxon B, ou de son lignage, va être artificiellement étendue et, sur cette durée, deux taxa seront comptabilisés (A + "B fantôme") au lieu d'un seul (A). La méthode cladistique postule que la relation ancêtre-descendant est indémontrable, et n'en tient donc pas compte, mais ça n'empêche pas qu'elle puisse parfois être inférée dans le registre fossile.
À l'inverse, un taxon fantôme peut recouvrir un grand nombre de taxa (inconnus mais ayant existé) : comptabiliser un taxon fantôme est donc une correction a minima. Par ailleurs, cette correction est asymétrique, puisque les dates d'extinction, elles, ne sont pas corrigées.
Ce type de corrections et la confrontation de la phylogénie obtenue avec un modèle de diversification à vitesse constante ont permis aux auteurs de proposer un schéma d'évolution de la diversité des Dinosauriens, comprenant plusieurs phases de diversification rapide, et de proposer des dates pour ces épisodes.
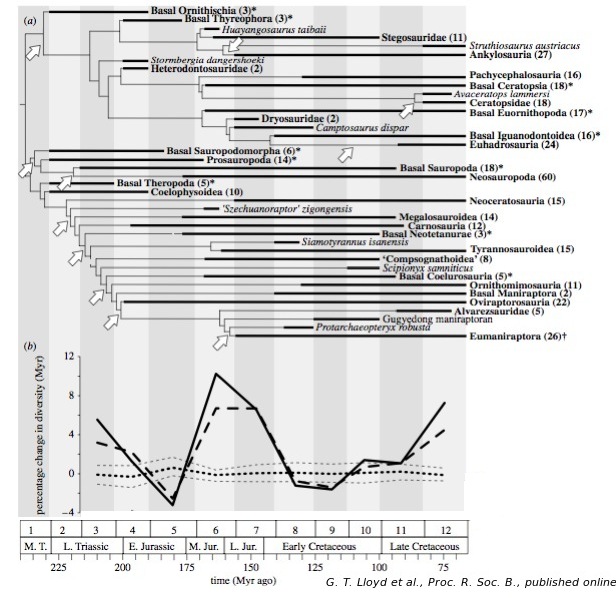
Source - © 2008 Lloyd et al., Proc. R. Soc. B., modifié
Figure 3. Évolution de la diversité des Dinosauriens
(a) Version résumée du super-arbre. Les 11 épisodes de diversification statistiquement significatifs sont marqués par des flèches blanches, indiquant les branches menant aux clades les plus diversifiés. Les taxa indiqués en gras représentent des regroupements de plusieurs grands clades, dont le nombre est mentionné entre parenthèses. Les astérisques (*) indiquent des regroupements formant un groupe paraphylétique et le symbole (†) un ensemble comprenant un clade actuel (à savoir les oiseaux). (b) Taux de diversification obtenu à partir de l'enregistrement fossile brut (ligne épaisse), sur le registre fossile associé à la prise en compte des "existences fantômes" (en tirets) et sur un traitement statistique par analyse de sous-échantillons (pointillés).
L'évolution des dinosauriens et l'environnement du Crétacé
Un intérêt de cette reconstitution de la biodiversité des dinosaures est qu'elle peut être comparée à l'évolution d'autres groupes d'organismes ou à des indicateurs paléo-environnementaux. Dans le cas des Dinosaures, une période intéressante est le Crétacé supérieur (vers 100 Ma). Au cours de cette période, d'importants changements des écosystèmes terrestres sont enregistrés : c'est l'explosion des Angiospermes (entre 125 et 80 Ma), mais aussi la diversification de nombreux clades d'Insectes, des Squamates (Lézards et Serpents), des Crocodiliens, des Oiseaux modernes... Cet épisode particulier de l'histoire de la vie terrestre aérienne est parfois nommé la « Révolution Terrestre du Crétacé » (KTR).
Or les précédentes estimations de la biodiversité dinosaurienne laissaient penser que ce groupe présentait lui aussi une diversification à ce moment. D'où l'hypothèse proposée d'un lien entre les diversifications des Dinosaures et des Angiospermes :
- certains grands groupes de dinosaures auraient pu sélectionner et favoriser les premières Angiospermes, dont les organes reproducteurs restaient près du sol et auraient ainsi pu échapper au broutage des dinosaures, ou au contraire en favorisant la dissémination des graines ;
- le développement des Angiospermes aurait, en retour, pu favoriser la diversification des dinosaures en leur fournissant de nouvelles sources de nourriture, donc de nouvelles niches écologiques.
Mais ce lien potentiel restait très critiqué, faute de preuve d'un net changement de régime alimentaire chez les dinosaures herbivores lors de la diversification des Angiospermes. Les résultats de Lloyd et al. continuent, à ce titre, de désavouer ces scénarios : selon eux, la rapide diversification des dinosaures au Campanien et au Maastrichtien serait un artefact que leur étude relativise largement. Ils obtiennent bien deux épisodes de diversification tardive à ce moment-là, correspondant au développement des Cératopsiens et des Hadrosaures, mais ils n'enregistrent pas d'accélération marquée de la diversification au Crétacé supérieur : l'essentiel de la diversification des Dinosaures se concentre au Trias et au Jurassique.
Toutefois, leur étude retrouve tout de même une diversification du groupe dans les derniers millions d'années du Crétacé, juste avant la crise K-T, les dinosauriens n'étaient donc nullement un groupe "fragile" ou "en déclin", ayant perdu de sa diversité par rapport au Jurassique, comme on a pu le soutenir autrefois, mais un groupe toujours dominant. L'impact de la crise K-T n'en paraît que plus important encore pour ce clade.
Ce premier exemple montre comment les méthodes d'analyse des données peuvent conduire à une remise en question des idées précédentes. Ces résultats sont importants et stimulants, mais ne referment pas le sujet pour autant : les techniques d'analyse employées étant encore en cours de développement et les données pouvant encore être améliorées.
La Paleobiology Database et les changements de biodiversité du Phanérozoïque
Le second exemple met l'accent sur l'importance de l'amélioration des bases de données et donc de l'augmentation du registre fossile, qui reste une nécessité. Il est surtout emblématique car il remet en question un travail précédent qui a fait autorité pendant des années.
Les courbes de biodiversité marine
La biodiversité marine du Phanérozoïque est mieux connue que la diversité terrestre, et la biodiversité des invertébrés marins mieux que celle des vertébrés du même milieu, les fossiles en étant plus nombreux, mieux préservés, et donc mieux documentés, etc. Depuis de nombreuses années, des chercheurs proposent des courbes de biodiversité (souvent au niveau du Genre ou de la Famille) issues de l'analyse des collections de fossiles marins disponibles dans les musées et les universités. Ils ont pu également en dériver des courbes de variation des taux d'apparition et de disparition de taxa. C'est par de tel travaux, sur les organismes marins et terrestres, qu'ont été démontré la réalité des épisodes d'extinction en masse et des fameuses cinq grandes "crises", qui se caractérisent justement par des taux d'extinction (à la fois en pourcentage de la diversité du moment et en nombre d'extinctions par unité de temps) largement supérieurs à ceux rencontrés avant ou après.
L'une des courbes les plus connues est celle obtenue par Jack J. Sepkoski (1948-1999) à partir du recensement de tous les genres d'invertébrés marins décrits depuis les débuts de la paléontologie scientifique (soit, à l'époque, la compilation de l'extension stratigraphique de plus de 31.000 genres appartenant à 4000 familles d'invertébrés à partir des données de la littérature). La courbe issue de ce colossal recensement (le "Compendium Sepkoski") fut reprise par de nombreux autres travaux ainsi que par les ouvrages de vulgarisation ou d'enseignement.
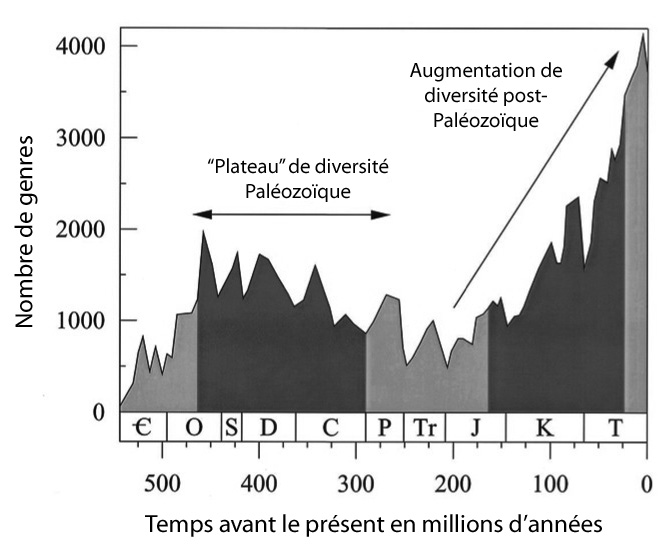
Source - © 2001 Newman, P.N.A.S. 98(11), modifié
Les intervalles en gris foncé correspondent aux périodes utilisées par Alroy et al. pour un première comparaison des résultats issus de la Paleobiology Database avec cette courbe classique (Alroy et al., PNAS 98(11), 2001).
Cette courbe fut évidemment critiquée, amendée et complétée par la suite : elle ne prenait pas en compte, initialement, des effets comme la relation entre biodiversité marine et surface des plateformes continentales ; des biais statistiques alors ignorés n'était pas corrigés, en particulier la différence entre "date de dernière apparition d'un fossile dans les roches" et "date réelle d'extinction", qui peut donner à une extinction géologiquement brutale l'aspect d'un changement progressif ("effet Signor-Lipps") ; l'identification de certains taxa a changé ; le registre fossile a été amélioré, etc.
Sur cette courbe classique, la biodiversité marine présentait une augmentation au début du Paléozoïque puis un plateau, suivi d'une augmentation continue et presque exponentielle à partir du Secondaire jusqu'à l'actuel. Mais cette expansion de la biodiversité pourrait n'être qu'un artefact lié à la représentation des roches. Plus il y a de roches fossilifères disponibles, plus on collecte de fossiles et plus on échantillonne de biodiversité. Les formations du Tertiaire affleurant plus et étant mieux préservées que les roches du Secondaire et celles-ci mieux que celles du Primaire, on peut s'attendre à une apparente augmentation de la biodiversité qui ne serait que le reflet de problèmes de préservations.
La meilleure manière de vérifier la fiabilité de ces résultats consiste évidemment à les retrouver à partir d'un nouveau corpus de données. C'est ce qu'a tenté l'équipe de John Alroy et al., en partant d'une nouvelle base de données, la Paleobiology Database.
La Paleobiology Database
La Paleobiology Database, développée depuis 2000, collecte et met à la disposition des chercheurs, via son site internet, le registre fossile de centaines de milliers de taxa (132.283 taxa, le 1er septembre 2008 à 21h, heure française). Et cet site, grâce à l'emploi d'outils informatiques modernes, ne se contente pas de rassembler les références taxinomiques et bibliographiques, il propose aussi au chercheur, ou plus généralement à l'internaute, toutes les informations récoltées sur chaque taxon : classification linnéenne, répartition géographique au cours du temps, données morphologiques de référence, etc. Il permet aussi de représenter "en ligne" la répartition d'un taxon fossile sur la carte paléogéographique d'une époque donnée. Bref un outil particulièrement puissant et "à jour".
Tout comme le "Compendium Sepkoski" a été complété par des taxa d'organismes terrestres (tétrapodes, insectes, plantes vasculaires...), la Paleobiology Database ne se limite absolument pas aux invertébrés marins. Mais comme cette catégorie de fossiles est, fondamentalement, la mieux fournie, avec des fossiles présents sur l'ensemble du Phanérozoïque, elle est la plus utilisable pour évaluer la diversité biologique avec des outils statistiques solides.
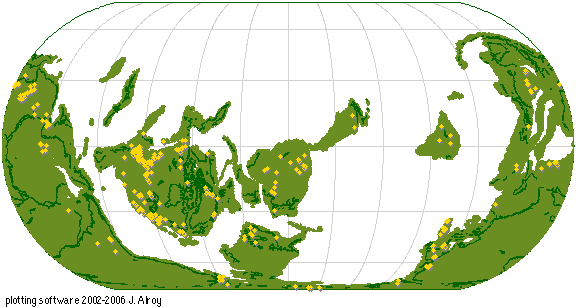
Source - © 2008 Paleobiology Database
Figure 5. Exemple de consultation de la Paleobiology Database
Positionnement sur un fond paléogéographique des Trilobites référencés dans la base de données pour la période du Cambrien.
La Paleobiology Database a donc été conçue, dès son lancement, avec pour objectif de décrire le registre fossile au plus près des données de terrain. Ainsi, un de ses avantages pour l'estimation de la biodiversité passée est qu'elle recense la présence d'un taxon fossile donné dans des collections précises, localisées géographiquement et temporellement. La base de Jack J. Sepkoski, elle, ne mentionnait que les dates d'apparition et de disparition de chaque taxon, sans préciser le nombre de spécimens et les différents sites de collecte du taxon pour cet intervalle de temps : un taxon rare et un taxon abondant se retrouvait donc sur un pied d'égalité dans le décompte de la biodiversité. Et, comme déjà dit auparavant, le biais lié aux conditions d'affleurement des roches n'était pas pris en compte.
Nouveaux résultats et révisions des idées
Avec cette nouvelle base, Alroy et al. ont pu procéder à une nouvelle estimation de la biodiversité des invertébrés marins au niveau taxinomique du Genre (plutôt que d'un groupe de rang supérieur, comme la Famille) au cours du Phanérozoïque, en appliquant des tests statistiques rigoureux.
- Les taxa ont été comptés en subdivisant le Phanérozoïque en intervalles de temps à peu près égaux (11 Ma).
- Un tirage au hasard a été effectué pour chaque intervalle de temps, jusqu'à obtenir un nombre identique (et important) de spécimens ; le nombre de Genres ainsi échantillonné dans chaque tranche temporelle a alors été compté et cette opération a été réitérée à de nombreuses reprises : une valeur moyenne de la diversité générique, assortie de barres d'erreur, pouvait alors être calculée ; cette opération permet de surmonter le problème du biais lié à "l'effort d'échantillonnage" (la plus ou moins bonne récolte de fossiles selon l'âge des terrains), puisque l'échantillonnage est ici fixé à la même valeur pour tous les intervalles.
- Une autre façon de corriger les biais d'échantillonnage consiste à tirer, pour chaque période, le même nombre de "collections" de fossiles (c'est-à-dire de sites fouillés), en prenant soin de tirer pour chaque intervalle de temps des collections dispersées géographiquement (afin de réduire le problème des différences de préservation des fossiles selon la géographie et les conditions climatiques).
- Les auteurs ont aussi choisi d'exclure de leur échantillonnage les sites et les spécimens les moins lithifiés et les coquilles fossiles ayant conservé leur aragonite originelle, donc les fossiles peu ou pas diagénétisés : ceci afin d'éviter les sites exceptionnels, ayant pu fournir des fossiles particulièrement fragiles. Les sites de ce genre sont en effet plus nombreux dans les terrains du Cénozoïque que dans ceux du Mésozoïque ou du Paléozoïque, leur prise en compte aurait introduit un biais en faveur de l'ère Tertiaire.
Conséquence immédiate de ces choix et de ces procédures, le nombre de genres obtenu par cette équipe (courbe ci-dessous) est systématiquement bien inférieur à ceux de la courbe de Sepkoski (ci-dessus). Mais c'est surtout la forme des deux courbes qu'il faut comparer.
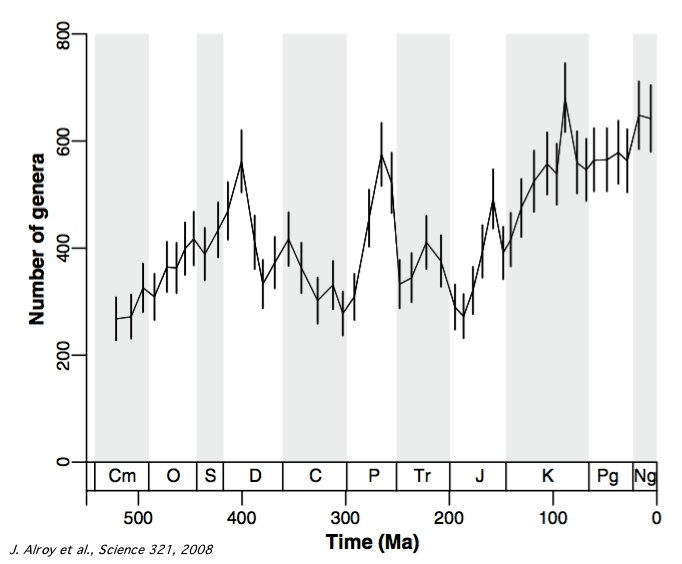
Source - © 2008 Alroy et al., Science 321, modifié
Chaque point représente la biodiversité des invertébrés marins sur un intervalle de temps d'environ 11 Ma. Les barres verticales délimitent l'intervalle de confiance à 95%.
Si certains points de la nouvelle courbe de la diversité des invertébrés marins publiée par cette équipe sont considérés comme douteux par les auteurs eux-mêmes (en particulier le petit pic du Crétacé supérieur), les grandes tendances paraissent robustes. « Plusieurs résultats-clés des courbes antérieures sont retrouvés : la radiation cambro-ordovicienne, la chute de 78% de la diversité à la fin du Permien, celle de 63% à la fin du Trias ».
Toutes les citations entre guillemets sont directement issues de l'article de d'Alroy et al.(Science 321, 2008)
Par contre, de nombreux résultats sont nouveaux. Le "« plateau »" du milieu du Paléozoïque est remplacée par "« une chute de la diversité au milieu du Dévonien qui n'est pas récupérée avant le Permien ». Qui plus est, ce déclin commence « bien avant la crise écologique du Frasnien-Famennien (fin du Dévonien, vers 365 Ma) et la diversité ne montre d'ailleurs pas de chute nette lors de cet épisode, ni lors de l'épisode glaciaire de la fin du Famennien, alors que la grande augmentation de la diversité au Permien, elle, coïncide, en gros, avec la déglaciation ». La récupération de diversité après la crise Permo-Trias paraît aussi particulièrement rapide et, « pour des raisons similaires », l'influence de la crise Crétacé-Tertiaire n'apparaît pas sur la courbe à cette résolution (c'est-à-dire avec ce pas d'échantillonnage de ~11 Ma). Ceci ne signifie pas que cette influence n'a pas existé, mais seulement qu'elle a été "épongée" très vite.
Autre différence importante par rapport aux résultats de Sepkoski, l'augmentation quasi-exponentielle de la diversité au Cénozoïque n'apparaît plus aussi clairement sur cette nouvelle courbe. « Les valeurs du Trias sont supérieures à celle du début du Jurassique, au lieu du contraire, et il y a peu de changement net entre le milieu du Crétacé (K) et le Paléogène (Pg) ». La biodiversité du Cénozoïque reste moins de deux fois plus importante que celle du Paléozoïque, alors que la courbe de Sepkoski présente une diversité récente plus de trois fois supérieure à celle du Primaire.
Avantage supplémentaire de cette nouvelle base de données : elle semble bien échantillonner les taxons de la zone tropicale, peut-être mieux que les précédentes bases, et elle permet, pour certains périodes au moins, d'évaluer les gradients latitudinaux de biodiversité (pour les invertébrés marins). Ainsi, pour le Néogène (du Miocène au Pléistocène), les auteurs retrouvent, dans leur échantillonnage, le fort gradient de diversité entre les tropiques et les hautes latitudes. Le tirage d'un même nombre de spécimens, soit dans le sous-ensemble des sites de basses latitudes, soit dans celui des sites de hautes latitudes, correspond à un nombre de genres bien plus élevé dans le premier cas que dans le second. La même procédure pour la période ordovicienne suggère que le gradient latitudinal de diversité était déjà présent à cette époque, bien que moins prononcé.
Enfin, pour obtenir une comparaison directe et quantifier l'influence des nouvelles méthodes sur la lecture de la biodiversité fossile, les auteurs ont réalisé une courbe de la biodiversité des métazoaires marins (excluant les tétrapodes) au cours des temps géologiques en comptabilisant les genres suivant la même procédure que J. J. Sepkoski. Les deux courbes se ressemblent alors beaucoup ce qui démontre que la Paleobiology Database offre une bonne description du registre paléontologique. Alroy et al. attribuent les différences entre la nouvelle courbe et celle de Sepkoski à la correction des biais analytiques. En particulier, la diversification exponentielle du Cénozoïque envisagée par de nombreux auteurs ne serait qu'un artefact. Ce qui pourrait impliquer que la biodiversité serait "contrainte", autrement dit ne pourrait peut-être pas augmenter indéfiniment comme elle paraît le faire sur la courbe de J. J. Sepkoski.
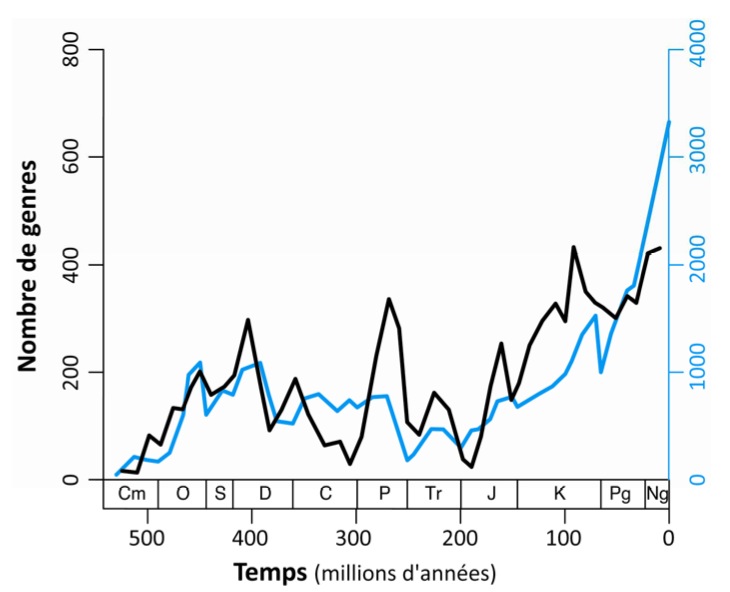
Source - © 2008 Loïc Villier
Les différences de calcul expliquent les différences d'échelle.
Conclusion
L'étude de la paléo-biodiversité, comme celle de la biodiversité actuelle, s'effectue aujourd'hui à travers des techniques de traitement des données pointues et récentes. Loin de n'être qu'une redite de travaux antérieurs, elle permet, en testant la solidité de résultats précédents, d'améliorer les connaissances et de reprendre des hypothèses et des idées plus anciennes. Elle est un préalable obligatoire à d'autres travaux, comme l'étude des grandes crises biologiques passées mais aussi la compréhension des mécanismes écologiques à l'origine de la biodiversité actuelle et récente (et donc des perturbations causées par l'activité humaine).
Ces deux études reposent sur la constitution de grandes bases de données, obtenues par la consultation de nombreuses références bibliographiques et sur la mise en commun du travail de nombreux chercheurs. Elles sont fondamentalement pluridisciplinaires, puisqu'elles font appel aux travaux de spécialistes de l'un ou l'autre des taxa considérés, de phylogénéticiens, de statisticiens et informaticiens. En paléontologie comme ailleurs, les programmes de recherches deviennent des travaux d'équipe, ou impose la coopération de plusieurs équipes.
Plus largement, ces travaux sont des illustrations de la modernité des recherches actuelles en paléontologie : l'image du paléontologue occupé à dégager à la brosse à dent un fossile encore inconnu, si elle n'est pas obsolète, loin s'en faut, n'est plus représentative d'une bonne part des travaux menés dans cette discipline. L'étude des organismes fossilisés, comme celle des organismes actuels, passe désormais, et de plus en plus, par la recherche in silico, ne serait-ce que pour pouvoir analyser efficacement l'énorme quantité d'information et de fossiles qu'ont récolté et que continuent à engranger les "paléontologues de terrain".
Références
- G.T. Lloyd, K.E. Davis, D. Pisani, J.E. Tarver, M. Ruta, M. Sakamoto, D.W.E. Hone, R. Jennings, M.J. Benton, 2008. Dinosaurs and the Cretaceous Terrestrial Revolution. Proceedings of the Royal Society B, 275, 1650, 2483-2490 - doi : 10.1098/rspb.2008.0715
- Communiqué de presse de l'Université de Bristol, 23 juillet 2008 : Dinosaurrific! New Dinosaur Supertree – the most comprehensive picture ever produced of how dinosaurs evolved
- O.R.P. Bininda-Edmonds, 2004. The evolution of supertrees.TRENDS in Ecology and Evolution, 19, 6, 315-322 - doi :10.1016/j.tree.2004.03.015
- A. Lane, C.M. Janis, J.J. Sepkoski, 2005. Estimating paleodiversities: a test of the taxic and phylogenetic methods. Paleobiology, 31, 1, 21-34 - doi :10.1666/0094-8373(2005)031<0021:EPATOT>2.0.CO;2
- J. Alroy et al., 2008. Phanerozoic Trends in the Global Diversity of Marine Invertebrates. Science, 321, 5885, 97-100 - doi :10.1126/science.1156963
- The Paleobiology Database

