Article | 09/06/2017
Homo naledi, la nouvelle star d'Afrique du Sud
09/06/2017
Résumé
Un lot de nouvelles données sur cet hominine décrit en 2015 vient d'être publié, et notamment une datation. Mais ces nouvelles analyses posent plus de questions sur le genre Homo qu'elles n'en résolvent.
En septembre 2015, l'équipe du paléoanthropologue Sud-africain Lee Berger publiait la découverte des restes de plusieurs squelettes d'un hominine jusque-là inconnu, attribué à une nouvelle espèce du genre Homo, baptisée Homo. naledi (en référence au nom du site en mangue locale [1]). Ce nouveau membre du genre humain présentait de nombreuses caractéristiques surprenantes, mais ne pouvait être précisément daté. En mai 2017, la même équipe publie, dans le même journal, un lot de résultats supplémentaires, comprenant la description d'autres restes et une proposition de datation. Ces données accentuent l'originalité de H. naledi, qui apparait simultanément assez archaïque dans sa morphologie et étonnamment récent.

Source - © 2015 D'après Berger et al. [1]
Figure 1. La présentation d'Homo naledi à la presse, en 2015
Un squelette reconstitué et une flopée d'os d'autres individus.
En 2015, quatre-vingt-dix ans tout juste après la publication par le paléontologue australien Raymond Dart (1893-1988), de la découverte en Afrique du Sud d'un fossile apparenté à l'Homme, Australopithecus africanus (« l'enfant de Taung »), les grottes karstiques de ce pays focalisent à nouveau l'attention des paléoanthropologues. Lee Berger, chercheur de l'université du Witwatersrand, présente aux médias un squelette étonnamment complet et de nombreux autres restes d'un nouvel hominine, que son équipe a baptisé Homo naledi (figure 1). À la place des seuls face et moulage endocrânien de l'« enfant » déterré par Dart, Berger annonce l'exhumation, depuis novembre 2013, de 1550 os appartenant à une quinzaine d'individus « au moins », et d'âges biologiques différents (jeunes et adultes) [1]. Par ailleurs, Berger et son équipe bouscule les habitudes par leurs méthodes de recherche et l'impact médiatique qu'ils donnent à leurs découvertes, ce qu'ils avaient déjà réussi pour un autre fossile, Australopithecus sediba, publié en 2010 dans la revue Science (cf. Australopithecus sediba, nouvel australopithèque d'Afrique du Sud). Cette fois, Lee Berger est allé encore plus loin [2] :
- Il finance l'exploration des réseaux karstiques du pays par des spéléologues chargés de signaler tout indice de fossile d'hominine ;
- pour accéder à la chambre où se trouvaient ceux-ci, il a engagé via internet de jeunes femmes paléontologues et spéléologues, suffisamment menues pour se glisser dans le boyau très étroit du système karstique qui permet d'y accéder (figure 3) ; le reste de l'équipe supervisait le dégagement des fossiles depuis l'extérieur, à l'aide de caméras ;
- il a également négocié une étroite collaboration avec le magazine National Geographic, qui finance les fouilles en échange d'un suivi exclusif de l'avancée des découverte ;
- enfin, l'équipe publie ses résultats dans une revue à comité de lecture mais en libre accès, apparue en 2012 et encore peu connue, eLife [3]. Bravade contre le système d'édition traditionnel ou refus des journaux les plus prestigieux de publier un fossile qui manque de l'information la plus cruciale, sa datation ? C'est en tout cas un choix que Berger et ses collaborateurs réitèrent en 2017.
Source - © 2017 D'après Google Earh, modifié
En 2015, ce nouvel hominine souffre en effet d'un manque majeur : son âge est inconnu. Ses caractéristiques anatomiques, elles, étonnent, de même que le contexte du site : il s'agit d'individus un peu plus grands que des australopithèques de plus de 2 millions d'années, mais à la capacité crânienne assez voisine (513 cm3 en moyenne, pour les individus adultes, contre 450 à 550 cm3) ; le squelette post-crânien (le squelette à partir de la première vertèbre) rappelle les australopithèques alors que le crâne évoque celui d'un Homo erectus (fort bourrelet sus-orbitaire, prognathisme net), une espèce datée de moins de 2 Ma, dont la capacité crânienne moyenne tourne plutôt autour de 1000 cm3 (figure 4).
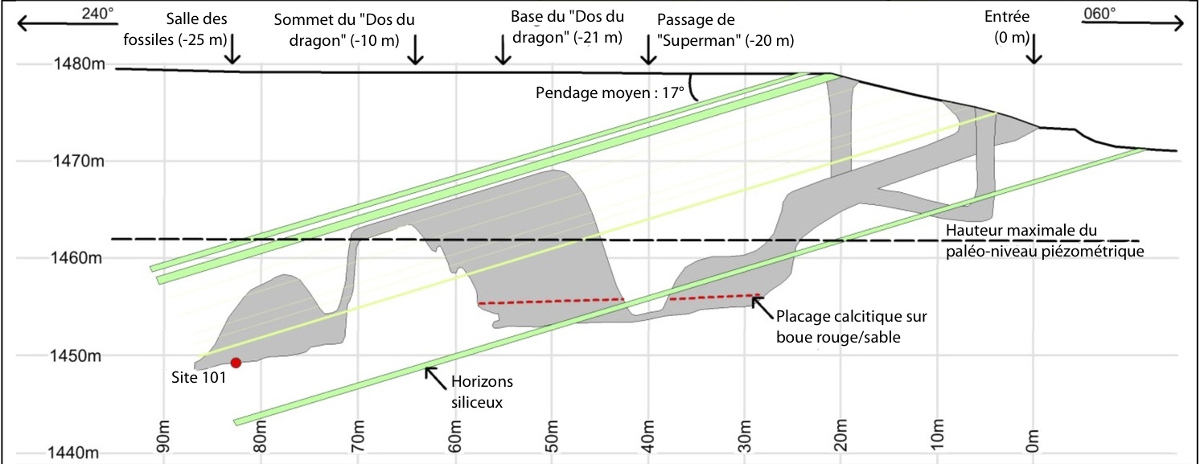
Source - © 2015 D'après Dirks et al. [12], francisé
Figure 3. Coupe du système de galerie de Rising Star montrant la voie d'accès aux fossiles publiés en 2015
Le « passage de Superman » n'est franchissable qu'en rampant dans le boyau, un bras tendu en avant, d'où son appellation.
Par ailleurs, l'explication la plus simple à la présence de squelettes de primates dans une cavité karstique Sud-africaine est celle d'un apport depuis l'extérieur, soit par des prédateurs, soit par une crue. Mais dans cette chambre-ci, les éléments squelettiques retrouvés semblaient tous parallèles les uns aux autres, ne portent pas de trace de charognage et ne sont pas associés à des ossements provenant d'autres animaux [2]. D'où la proposition d'une disposition intentionnelle des squelettes, donc d'une forme d'inhumation de ces quinze individus par leurs congénères. Que la cavité soit très difficile d'accès (figure 3), même pour des individus de petite taille, rend encore plus énigmatique cette accumulation de squelettes : elle nécessiterait un transport très compliqué, qui plus est dans l'obscurité la plus complète. On peut soupçonner que la grotte possédait autrefois une autre ouverture, plus large et plus accessible.

Source - © 2008 D'après Robson & Wood [13], modifié
En mai 2017, Lee Berger et ses associés publient trois nouveaux articles, toujours dans eLife. L'un d'eux décrit de nouveaux éléments squelettiques découverts dans une seconde chambre du réseau karstique et attribués à la même espèce [4]. Ils comprennent un squelette partiel, un crâne fragmentaire auquel manque une partie de la face, mais dont le volume a pu être reconstitué (610 cm3), et sa mandibule. Un deuxième article [5] propose une longue réponse (59 pages !) à la question cruciale de l'âge de ces fossiles. L'équipe a croisé les résultats de plusieurs techniques de datation sophistiquées, développées notamment en Australie pour dater justement les sites archéologiques dans des environnements calcaires et désertiques. Certaines, comme la luminescence stimulée optiquement (Optically Stimulated Luminescence, OSL) et la datation uranium-thorium (déséquilibre des chaines radioactives de l'uranium) ont été appliquées aux placages calcitiques (flowstones) de la grotte ; d'autres ont pu être utilisées directement sur quelques dents des spécimens (datation par résonance de spin électronique, Electronic Spin Resonance [6]). Les résultats obtenus ajoutent à la confusion : ces fossiles auraient entre 236 000 et 335 000 ans ! C'est-à-dire qu'ils dateraient de la fin du Pléistocène moyen (781 000 à 126 000 ans) et seraient contemporains des néandertaliens d'Eurasie et des dénisoviens d'Asie. Cette découverte d'individus à « petite tête » et pourtant récents rappelle évidemment le cas des « petits Hommes » de Florès (Homo floresiensis), dont des résultats récents ont réévalué l'âge entre 50 000 et 190 000 ans [7] (au lieu des 18 000 ans attribués au principal squelette lors de sa publication en 2003 et des 12 000 à 95 000 ans avancés pour l'ensemble des pièces). Dans ce dernier cas, les erreurs de datation provenaient en partie d'une corrélation erronée entre les restes fossiles et les couches sédimentaires (certaines pièces du squelette fragmenté se trouvaient inclus dans des couches plus récentes). Dans le cas d'Homo naledi, la concordance des datations effectuées sur les sédiments et sur les dents elles-mêmes semble exclure ce type d'erreur.
Qu'en conclure, pour l'instant ? Les dates récentes de ce nouvel hominine étonnent parce que son anatomie faisait attendre a priori une date beaucoup plus ancienne, supérieure à 1 million d'années, qui l'aurait rapproché d'hominines au cerveau un peu supérieur, comme Homo erectus ou Homo ergaster, ou des espèces encore plus anciennes, comme Homo habilis ou les derniers Australopithèques (figure 4). Cela n'aurait pas contredit la tendance à l'augmentation du volume cérébral observé dans le genre Homo depuis 1,8 Ma avec Homo erectus et qui parait s'accentuer à partir de 800 000 ans (figure 4).
Si l'attribution de ces fossiles au genre Homo est confirmée, il faut en conclure que ce genre a été plus diversifié encore qu'on ne le pensait jusqu'à présent, avec plusieurs branches dont certaines seulement auraient développé un grand cerveau. Cela invaliderait au passage l'une des hypothèses explicatives à cette tendance évolutive, celle du lien entre acquisition de la bipédie et développement cérébral. Cela donne par contre un relief supplémentaire à une publication récente de paléogénétique co-signée par le désormais célèbre paléogénéticien suédois Svaante Pääbo, qui montre qu'une mutation génétique propre à l'Homme, et dont l'apparition remonterait à Homo erectus, aurait permis ce développement cérébral particulier. Dans un gène humain lui-même issu de la duplication d'un autre gène préexistant, une mutation unique (une transversion C → G) induit la coupure d'une petite séquence de l'ARN messager produit par transcription de ce gène. Cette modification ôte à ce gène sa capacité à produire une protéine, mais contribue à accentuer la prolifération des cellules progénitrices des neurones du néocortex [8]. Elle pourrait donc avoir contribué à cette tendance évolutive si particulière vers un cerveau « anormalement » volumineux chez un mammifère de cette taille. D'où l'empressement de la presse de vulgarisation scientifique même la plus fiable à y voir « le moment où nous sommes devenus humains », comme l'a titré un contributeur de la revue Pour la Science sur le site de la revue [9] avant de réaménager son article dans le numéro de mars 2017 du journal sous le titre plus sobre de [la] mutation génétique qui a profité au cerveau d'Homo sapiens [10].
Peut-être Homo naledi ne descendait-il pas du groupe humain dans lequel s'est fixée cette mutation ? Mais attribuer à un seul changement génétique l'augmentation du volume cérébral qui s'est déroulée sur plus de 800 000 ans dans le genre Homo semble simpliste, et d'autres théories explicatives, compatibles avec celle-ci, restent envisageables, comme le développement des liens sociaux et l'amélioration de la qualité de l'alimentation, qui auraient pu initier des boucles de rétroaction positives avec l'amélioration des capacités cognitives. Quoi qu'il en soit, il semblerait que les formes fossiles aujourd'hui rassemblées dans le genre Homo aient été plus variées génétiquement, anatomiquement et morphologiquement, qu'on ne le pensait jusqu'ici. Ce qui revient aussi, pour les systématiciens, à un problème plus difficile à trancher : cette diversité n'est-elle pas trop grande pour un niveau taxinomique aussi bas que le genre ? Faudrait-il limiter le genre Homo aux bipèdes à gros cerveau, et en créer un autre pour les formes « à petite tête » ? Ou au contraire la limitation actuelle du genre Homo à une seule espèce, la nôtre, nous induit-elle à restreindre à l'excès sa définition ? Là encore, le problème n'est pas tout à fait nouveau : de plus en plus, aujourd'hui, certains paléoanthropologues rechignent à inclure dans le genre Homo ses tous premiers représentants officiels, Homo habilis et Homo rudolfensis, dont l'anatomie reste très proche de celles des australopithèques [11]. Pour eux, le genre Homo ne commencerait vraiment qu'avec les Homo erectus et Homo ergaster, nettement plus grands et au cerveau sensiblement plus volumineux (figure 4). Ce qui pose la question des relations phylogénétiques d'Homo naledi avec les autres Homo : est-il plus apparenté aux anciens Homo habilis qu'aux Homo erectus ou à d'autres formes plus récentes ?
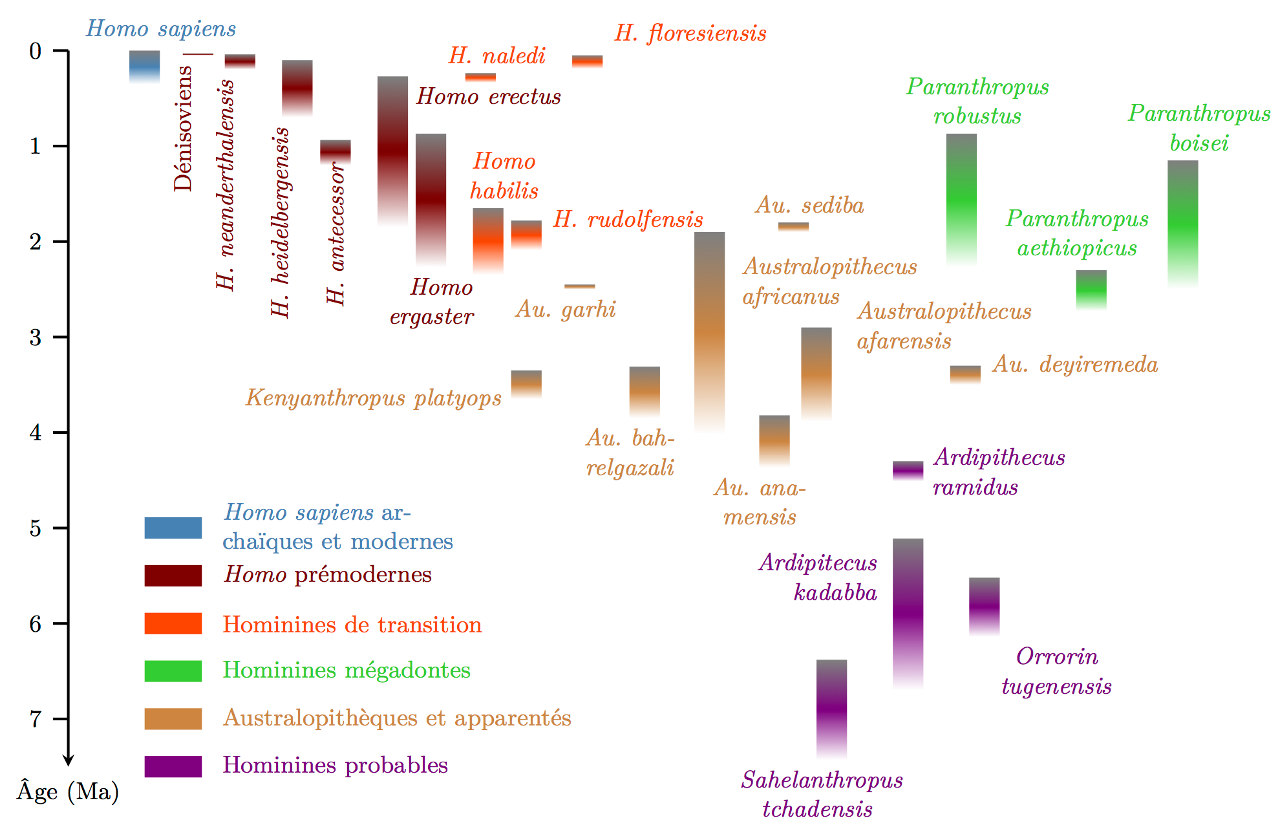
Source - © 2017 Cyril Langlois, inspiré de B. Wood & E. Boyle, 2016 [11]
Les « hominines de transition » désignent, ici, tous les taxons placés dans le genre Homo mais de petite capacité crânienne (< 800 cm3).
Ce qui parait certain, c'est que Lee Berger, en dépit des critiques sur sa méthode qui fait de la recherche paléoanthropologique une quasi-entreprise et exploite sans état d'âme l'aura médiatique dont elle bénéficie, vient rappeler l'importance de l'Afrique du Sud dans l'évolution africaine du rameau humain, à côté de la vallée du Rift (Éthiopie, Tanzanie, Kenya) et du Tchad. Et démontre que le tableau actuel, déjà bien compliqué (figure 5), reste fragmentaire.
Bibliographie
[1] L.R. Berger, J. Hawks, D.J. de Ruiter, S.E. Churchill, P. Schmid, L.K. Delezene, T.L. Kivell, H.M. Garvin, S.A. Williams, J.M. DeSilva, M.M. Skinner, C.M. Musiba, N. Cameron, T.W. Holliday, W. Harcourt-Smith, R.R. Ackermann, M. Bastir, B. Bogin, D. Bolter, J. Brophy, Z.D. Cofran, K.A. Congdon, A.S. Deane, M. Dembo, M. Drapeau, M.C. Elliott, E.M. Feuerriegel, D. Garcia-Martinez, D.J. Green, A. Gurtov, J.D. Irish, A. Kruger, M.F. Laird, D. Marchi, M.R. Meyer, S. Nalla, E.W. Negash, C.M. Orr, D. Radovcic, L. Schroeder, J.E. Scott, Z. Throckmorton, M.W. Tocheri, C. VanSickle, C.S. Walker, P. Wei, B. Zipfel, 2015. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa, eLife, 4:e09560, [doi:10.7554/elife.09560]
[2] K. Wong, 2017. L'incroyable Homo naledi, Dossiers Pour la Science, vol. 94, p. 39‑45
[3] eLife, disponible sur https://elifesciences.org [consulté le 07 juin 2017]
[4] J. Hawks, M. Elliott, P. Schmid, S.E. Churchill, D.J. de Ruiter, E.M. Roberts, H. Hilbert-Wolf, H.M. Garvin, S.A. Williams, L.K. Delezene, E.M. Feuerriegel, P. Randolph-Quinney, T.L. Kivell, M.F. Laird, G. Tawane, J.M. DeSilva, S.E. Bailey, J.K. Brophy, M.R. Meyer, M.M. Skinner, M.W. Tocheri, C. VanSickle, C. S. Walker, T.L. Campbell, B. Kuhn, A. Kruger, S. Tucker,A . Gurtov, N. Hlophe, R. Hunter, H. Morris, B. Peixotto, M. Ramalepa, D. van Rooyen, M. Tsikoane, P. Boshoff, P.H.G.M. Dirks, L.R. Berger, 2017. New fossil remains of Homo naledi from the Lesedi Chamber, South Africa », eLife, 6:e24232,[doi:10.7554/eLife.24232]
[5] P.H.G.M. Dirks, E.M. Roberts, H. Hilbert-Wolf, J.D. Kramers, J. Hawks, A. Dosseto, M. Duval, M. Elliott, M. Evans, R. Grün, J. Hellstrom, A.I.R. Herries, R. Joannes-Boyau, T.V. Makhubela, C.J. Placzek, J. Robbins, C. Spandler, J. Wiersma, J. Woodhead, L.R. Berger, 2017. The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa, eLife, 6:e24231, [ doi:10.7554/eLife.24231]
[6] J.-J. Bahain, 2007. La méthode de datation par résonance de spin électronique (ESR) au Muséum national d'histoire naturelle. Vingt ans de recherches méthodologiques et d'applications géochronologiques, Habilitation à diriger des recherches, Univ. Michel de Montaigne - Bordeaux III
[7] T. Sutikna, M.W. Tocheri, M.J. Morwood, E. Wahyu Saptomo, Jatmiko, R. Due Awe, S. Wasisto, K.E. Westaway, M. Aubert, B. Li, J.-X. Zhao, M. Storey, B.V. Alloway, M.W. Morley, H.J.M. Meijer, G.D. van den Bergh, R. Grün, A. Dosseto, A. Brumm, W.L. Jungers, R.G. Roberts, 2016. Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia, Nature, 532, 7599, 366‑369, [doi:10.1038/nature17179]
[8] M. Florio, T. Namba, S. Pääbo, M. Hiller, W. B. Huttner, 2016. A single splice site mutation in human-specific ARHGAP11B causes basal progenitor amplification, Sci. Adv., 2,12, e1601941, [doi:10.1126/sciadv.1601941]
[9] S. Bohler, 2017. Le moment où nous sommes devenus humains, Pour la Science [En ligne, consulté le 07 juin 2017]
[10] S. Bohler, 2017. Une mutation génétique qui a profité au cerveau d'Homo sapiens, Pour la Science, 473, 8
[11] B. Wood E.K. Boyle, 2016. Hominin taxic diversity: Fact or fantasy?, Am. J. Phys. Anthropol., 159, S61, 37–78, [doi: 10.1002/ajpa.22902]
[12] P.H.G.M. Dirks, L.R. Berger, E.M. Roberts, J.D. Kramers, J. Hawks, P.S. Randolph-Quinney, M. Elliott, C.M. Musiba, S.E. Churchill, D.J. de Ruiter, P. Schmid, L.R. Backwell, G.A. Belyanin, P. Boshoff, K.L. Hunter, E.M. Feuerriegel, A. Gurtov, J. du G. Harrison, R. Hunter, A. Kruger, H. Morris, T.V. Makhubela, B. Peixotto, S. Tucker, 2015. Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa, eLife, 4:e09561, [doi:10.7554/eLife.09561]
[13] S.L. Robson, B. Wood, 2008. Hominin life history: reconstruction and evolution, Journal of Anatomy, 212, 94‑425, [doi:10.1111/j.1469-7580.2008.00867.x]