
Article | 28/04/2010
Plumes fossiles et évolution des oiseaux : quelques nouveautés
28/04/2010
Résumé
De récentes découvertes de fossiles permettent de rediscuter la phylogénie des dinosaures et, entre autres, la place des oiseaux. L'étude de dinosaures à plumes comparés aux oiseaux actuels aboutit à redéfinir la spécificité des oiseaux mais aussi à remonter à un aspect longtemps jugé inaccessible : la couleur des plumes fossiles.
Table des matières
Introduction
L'automne 2009 et le début d'année 2010 ont été riches en nouveautés dans le domaine de la paléontologie des oiseaux, avec une nouvelle illustration d'une "course à la publication" sur un thème très médiatique.
Dans son numéro du 1er octobre 2009, la revue britannique Nature exposait l'analyse d'un nouveau fossile de dinosaure « emplumé », Anchiornis huxleii [1], extrait, comme la plupart des précédents, d'un site fossilifère de la province du Liaoning, en Chine. Cette découverte pourrait presque passer pour banale, si cet animal ne venait pas compléter le registre fossile du Jurassique supérieur pour lequel on ne disposait jusqu'ici que de fossiles d'oiseaux (en particulier Archæopteryx), mais pas de représentants des groupes de dinosaures emplumés phylogénétiquement proches des oiseaux [2].
Quelques mois après, l'autre grande revue scientifique Science propose la description d'un nouveau dinosaure emplumé mais non-volant [3], qui vient à son tour éclairer l'histoire jurassique d'un des groupes de dinosaures les plus proches des oiseaux.
Début mars 2010, enfin — après un communiqué de presse paru dès le 27 janvier 2010— Nature publie une étude sino-britannique qui, à partir de l'examen méticuleux de fossiles d'oiseaux et de dinosaures emplumés, affirme pouvoir en reconstituer une caractéristique généralement considérée comme à jamais inaccessible sur les fossiles : la couleur [4]. Et quelques jours plus tard, c'est de nouveau au tour de Science d'annoncer la reconstitution complète, par une autre équipe (Li et al., Science Express Report, 4 février 2010), des couleurs... d'Anchiornis [5], le fossile décrit quelques mois plus tôt dans les pages de son concurrent.
Plumes, dinosaures et oiseaux : état des connaissances
Les relations dinosaures / oiseaux
L'inclusion des oiseaux (Aves) dans le clade des dinosaures est aujourd'hui établi et accepté par la majorité des paléontologues, sur la base de données anatomiques et, évidemment, des découvertes récentes — en provenance de Chine pour la plupart — de nombreux fossiles de dinosaures non-volants et pourtant emplumés.
Les dinosaures au sens classique, sans les oiseaux, constituent donc un groupe paraphylétique1, sans signification évolutive. Le nom de Dinosauriens (Dinosauria) désigne désormais le groupe monophylétique comprenant les dinosaures « classiques » et les oiseaux.
Comment définir alors un oiseau, puisque la plume n'en est plus le caractère-diagnostic ? L'idée actuellement retenue est que leur caractéristique fondamentale n'est plus la plume, mais le vol (battu, actif et non simplement plané) [6] même si celui-ci peut secondairement avoir été perdu, totalement ou partiellement, comme chez les autruches ou les poules.
Les analyses cladistiques actuelles non seulement lient les oiseaux aux dinosaures, mais les incluent parmi les multiples groupes composant le clade des Dinosauriens : les oiseaux appartiennent au grand clade des dinosaures saurischiens2, parmi ceux-ci à l'ensemble des théropodes (carnivores bipèdes) et, parmi les nombreux sous-clades qui le compose, au groupe des Maniraptora. Au sein des Maniraptora, tous les oiseaux depuis l'Archæopteryx sont rassemblés dans le clade des Avialea qui forme, avec ses deux groupes-frères, les Troodontidae et les Dromæosauridae, le clade des Paraves (les « presque oiseaux ») (cf. figure 1).
Les plus proches parents des oiseaux parmi les Maniraptora étaient de petits dinosaures bipèdes, dont beaucoup arboraient des plumes ou des filaments tégumentaires semblables aux plumes de duvet des jeunes oiseaux actuels, ou encore une association de plusieurs types de plumes.
Pour Kevin Padian et Armand de Ricqlès [6], les oiseaux descendent donc de dinosaures caractérisés non pas seulement par leurs plumes, mais surtout par une petite taille et des taux de croissance particulièrement élevés. L'étude de la microstructure des os, y compris fossiles, permet en effet d'estimer la vitesse de croissance de l'animal. Cette méthode suggérait déjà que les dinosaures dans leur ensemble se signalaient par une croissance très rapide, mais cette particularité s'est encore accentuée chez les oiseaux et leurs plus proches parents dinosauriens.
1 Un groupe est dit monophylétique s'il comprend un ancêtre et tous ses descendants, paraphylétique s'il contient un ancêtre et une partie seulement de ses descendants.
2 Dinosaures à "bassin de reptile" et non à "bassin d'oiseau" comme l'autre grand groupe des Ornitischiens, bel exemple de convergence évolutive.
La plume chez les dinosaures
Le registre fossile des dinosaures emplumés s'est développé avec les fouilles menées dans les formations du Crétacé inférieur de la province du Liaoning, en Chine. Ces roches ont livrés plusieurs fossiles de dinosaures et d'oiseaux sur lesquels ont été identifiés, dans des groupes distincts, différents types de plumes qui, pour certaines, rappellent différents stades de développement des plumes des oiseaux actuels, alors que d'autres présentent une structure originale. On en conclut que l'apparition, et la diversification, des plumes s'est effectuée parmi les dinosaures théropodes : elle pourrait être apparue chez l'ancêtre commun à tous les Théropodes cœlurosaures (cf. figure 1).
La plume ne peut donc plus être considérée comme une adaptation liée au vol. Son utilisation dans le vol constitue un "changement de fonction" d'une structure pré-existante, ce que Stephen Jay Gould et Elizabeth Vrba ont proposé d'appeler une « exaptation ». La fonction originelle des premières plumes est donc autre, et sans lien avec le vol.
Quelle a pu être, alors, la fonction des premières plumes ? L'hypothèse privilégiée aujourd'hui relie plusieurs éléments :
- les données histologiques et isotopiques en faveur d'un métabolisme actif et maintenu, donc de l'homéothermie (le « sang chaud »), chez les dinosaures théropodes ;
- la petite taille générale des groupes de dinosaures à plumes les plus basaux et des oiseaux anciens.
Ces caractéristiques impliquent, comme pour les Mammifères, que ces petits animaux, au rapport surface/volume élevé, perdaient rapidement leur chaleur, ce qui leur imposait un métabolisme très actif pour maintenir leur température corporelle. La plume de type duvet, comme le poil des Mammifères, serait alors une réponse adaptative à cette déperdition de chaleur en permettant le développement d'une couche d'air isolante.
Chez les dinosaures porteurs de plumes plus longues, ces dernières avaient probablement d'autres fonctions. Diverses hypothèses sont évoquées, par exemple un rôle de caractère sexuel, qu'il est difficile de démontrer. Chez certains de ces dinosaures bipèdes actifs, coureurs, des études biomécaniques suggèrent que les longues rémiges présentes sur les bras et la queue pouvaient peut-être contribuer à améliorer la stabilité de la course ou l'équilibre lors de brusques changements de direction. Ces animaux arboraient en effet de longues plumes asymétriques, semblables à celles des ailes des oiseaux. Ce type de plume n'est donc pas, lui non plus, un apanage des oiseaux en lien avec le vol. En effet, comme le signalent Kevin Padian et Armand de Ricqles [6], « les plumes asymétriques n'améliorent ni le vol, ni même la portance ; elles réduisent la traînée, ce qui est utile pour tout animal se déplaçant à une vitesse élevée ».
Si toutes ces interprétations semblent constituer un ensemble cohérent, l'histoire évolutive du clade des théropodes et les relations exactes entre les anciens oiseaux et les groupes de dinosaures phylogénétiquement les plus proches d'eux, sont encore loin d'être complètement élucidées.
Les premiers fossiles reconnus comme des oiseaux au sens strict, pourvus de plumes et anatomiquement capables d'un vol battu, datent du Jurassique récent, avec évidemment les dix exemplaires disponibles à ce jour du fameux Archæopteryx. Par contre, les fossiles représentatifs des taxa les plus proches phylogénétiquement des oiseaux (Alvarezsauridés, Troodontidés, Dromæosauridés) sont pour la plupart d'âge crétacé, donc plus récents qu'Archæopteryx. Certains de ces groupes pourraient effectivement être plus récents que la lignée des oiseaux (Troodontidés et Dromæosauridés ; cf. figure 1) mais d'autres, comme les Alvarezsauridés, seraient aussi anciens, ou plus anciens, que les oiseaux (cf. figure 1). Faute de fossiles du Crétacé inférieur et du Jurassique supérieur, il faut leur supposer une "lignée fantôme" (ghost lineage) remontant jusqu'à leur ancêtre commun avec les oiseaux.
Enfin, les convergences et les parallélismes évolutifs (homoplasies) entre les fossiles de Maniraptoriens sont nombreuses et compliquent le positionnement précis des oiseaux au sein de ce groupe [6].
C'est sur cette histoire ancienne des groupes de dinosaures à plumes non-aviens que les fossiles récemment publiés apportent de nouveaux éclairages... assortis de nouvelles questions.
Le registre fossile jurassique des dinosaures
Haplocheirus sollers, l'alvarezsauroïdé jurassique
Le fossile décrit en mars 2010 par Choiniere et al. [3], provenant du bassin de Junggar, dans le Xinjiang, à l'extrémité occidentale de la Chine, ajoute un nouveau représentant au groupe encore assez mystérieux des Alvarezsauroidea, l'un des groupes-frères putatifs du clade des Paraves (cf. figure 1).
Ce fossile, dénommé Haplocheirus sollers, vient compléter la liste des représentants de ce clade encore peu connu. Mais surtout, l'âge de ce fossile a été encadré entre deux datations radiométriques de 158,7 ± 0,3 et 161,2 ± 0,2 Ma, autrement dit à l'Oxfordien. Jusqu'ici, les fossiles rangés dans ce groupe des Alvarezsauroidea (cf. figure 1) n'avaient été exhumés que dans des formations du Crétacé supérieur d'Amérique du Nord, d'Amérique du Sud, d'Asie et d'Europe.
Haplocheirus sollers démontre donc que cette lignée remonte bien, comme le suggéraient les phylogénies, au Jurassique moyen ou supérieur. Puisque l'apparition de ce clade est forcément encore antérieur à ce fossile, il faut bien lui supposer un "lignage fantôme" d'au moins de 63 millions d'années ! Par ailleurs, cette découverte suggère que ce groupe serait apparu et aurait entamé sa diversification en Asie.
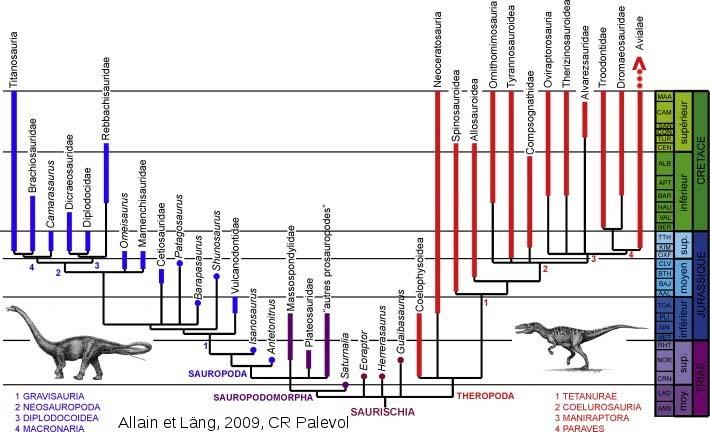
Source - © 2009 R. Allain et É. Läng, Comptes Rendus Palevol [7]
Les oiseaux (Archæopteryx et autres oiseaux fossiles) apparaissent sous l'appellation Avialae, en haut à droite.
Mais, surtout, Haplocheirus est plus ancien (d'une quinzaine de millions d'années) que le premier représentant attitré des Aviales, Archæopteryx. Cette découverte soutient donc l'une des hypothèses phylogénétiques proposées, qui place le branchement des Alvarezsauroidea avant la diversification du clade des Paraves (Avialae + Troodontidae + Dromæosauridae).
Il confirme également que les Alvarezsauroïdés, en particulier les formes dérivées du Crétacé, présentaient de nombreuses convergences évolutives avec les oiseaux. L'une des plus évidentes est la miniaturisation : Haplocheirus, dont le fossile est décrit comme « quasi-complet »,est à ce jour le plus grand des Alvarezsauroidea (avec une longueur totale estimée entre 1,90 m et 2,30 m). Or la réduction de taille, selon les auteurs de l'article, est une tendance évolutive rare chez les dinosaures, et justement remarquable chez les oiseaux.
Par ailleurs, ce premier représentant des Alvarezsauroïdés montre déjà certains traits typiques des espèces plus tardives de ce clade, en particulier une main au nombre de doigts réduit. Néanmoins, chez celui-ci, la main était encore fonctionnelle, c'est-à-dire mobile et capable de s'agripper, capacité perdue chez les formes crétacées, chez qui le membre supérieur était atrophié et les doigts réduits et pour certains fusionnés... une évolution là encore semblable à celle des oiseaux.
Anchiornis huxleyi, vieux et des plumes aux pattes
L'autre animal tout récemment décrit, là encore à partir d'un fossile presque complet et toujours issu des gisements chinois [1], appartient à un groupe encore plus proche des oiseaux, placé au sein même, cette fois, du clade des Aviales, les Troodontidés.
Lui aussi est d'abord intéressant par son âge : les sédiments desquels il a été extrait (la formation Tiaojishan, dans le district du Jianchang, province du Liaoning, au Nord-Est de la Chine, cette fois) ont été datés entre 161 et 151 Ma, soit, là encore, légèrement plus vieux que les calcaires de Solnhofen d'où provient le premier spécimen d'Archæopteryx (écart de 5 à 10 Ma). Et, là encore, les Troodontidés connus jusque là étaient tous bien plus récents.
Cet âge a encore d'autres conséquences. Selon la phylogénie, les Troodontidés formeraient un clade monophylétique avec les Dromæosauridés, groupe-frère des Aviales. Ce rapprochement n'est pas assuré, et plusieurs auteurs se bornent à indiquer une « trichotomie » (division en trois branches) entre Aviales, Troodontidés et Dromæosauridés. Quoi qu'il en soit, si Anchiornis est un Troodontidé, cela impose que la divergence Troodontidés-Dromæosauridés est déjà effective, donc que l'ancêtre commun des deux groupes est plus ancien. Ce qui repousse également l'ancêtre commun des trois groupes Troodontidés, Dromæosauridés et Aviales, c'est-à-dire l'ancêtre commun à l'ensemble monophylétique des Paraves. L'apparition et la diversification de ce clade remonteraient alors au Jurassique moyen.
Anchiornis est donc relativement proche de cet ancêtre commun des Paraves. On peut alors supposer, si l'on suit toujours la phylogénie de la figure 1, qu'il possède, à côté de ses caractéristiques spécifiques (autapomorphies) et des spécificités du clade (Troodontidés + Dromæosauridés), leurs synapomorphies, un certain nombre de caractères plésiomorphes, éventuellement perdus ou modifiés chez les représentants plus récents de chaque clade, mais présents et partagés par les plus anciennes formes. Par exemple, Anchiornis avait des membres supérieurs dont la longueur relative était supérieure à celle des Troodontidés plus récents. Mais cette longueur relative est semblable à celle des Aviales, ce qui complique d'autant son positionnement phylogénétique.
Par ailleurs, une particularité d'Anchiornis huxleyi est de montrer des plumes de grande taille non seulement sur les membres antérieurs et sur une longue queue, mais également sur les pattes postérieures (cf. figure 2b). Cette configuration n'est pas totalement surprenante, car elle avait déjà été découverte auparavant, mais sur un Dromæosauridé du Crétacé, Microraptor gui d'abord, puis sur un autre spécimen, Pedopenna daohugouensis, peut-être légèrement antérieur à Archæopteryx [9], et qui pourrait appartenir aux Aviales, donc aux oiseaux.

Source - © 2009 Hu et al., Nature [1]
Source - © 2010 Chuang Zhao & Lida Xing
Figure 2. Le fossile d'Anchiornis huxleyi et sa reconstitution proposée en octobre 2009
(a) Le fossile publié par Nature [1]. La barre d'échelle représente 5 cm.
(b) Reconstitution initiale d'Anchiornis.
On doit par conséquent en conclure que :
- cet aspect à quatre ailes n'est pas une originalité de Microraptor, comme on l'a pensé initialement ;
- il est en réalité partagé avec les Troodontidés et peut-être les premiers Aviales (si Pedopenna en est un), donc probablement présent chez l'ancêtre commun aux trois groupes.
L'interprétation évolutive la plus parcimonieuse de tout cela est que la configuration à quatre ailes représente l'état primitif des Paraves et non un bizarre état dérivé des groupes de « presque oiseaux », et que les oiseaux ont perdu par la suite cet empennage postérieur.
Remarquons ici que Microraptor gui a fait l'objet d'une reconstruction anatomique et d'une étude aérodynamique [8] à partir desquelles les auteurs ont conclu que cet animal pouvait effectuer un vol plané, en se lançant d'un arbre, par exemple, en plaçant les pattes arrières sous le corps et en étalant leurs plumes dans un plan parallèle à celui des ailes antérieures. Microraptor aurait donc plané comme un avion biplan du début du XXème siècle (cf. figure 3). Anchiornis était-il lui aussi capable de planer ? Les auteurs ne le précisent pas, mais les plumes visibles sur les pattes postérieures paraissent beaucoup plus courtes que celles de Microraptor.
![La position de vol plané en « biplan » de Microraptor gui, proposée par S. Chatterjee et R. J. Templin [8]](https://planet-terre.ens-lyon.fr/planetterre/objets/Images/plumes-dinosaures-oiseaux/vol-Microraptor.png)
Source - © 2007 Chatterjee & Templin, modifié [8]
La barre noire représente 5 cm.
Mélanosomes fossiles et couleurs des vieilles plumes
À côté de cet aspect phylogénétique, l'autre actualité paléontologique des dinosaures emplumés porte sur leur aspect extérieur : deux équipes viennent de proposer une reconstitution des couleurs de ces animaux.
Les couleurs de Sinosauropteryx et Confuciusornis
La première de ces publications [4] s'est penché sur trois fossiles :
ceux de deux dinosaures théropodes :
- Sinosauropteryx, placé parmi les Compsognathidae (cf. figures 1 et 6a) et apparemment recouvert d'un "pelage" de filaments, interprétés par certains comme des plumes très simples, mais par d'autre comme des traces de collagène dégradé ;
- Sinornithosaurus, un dromæosaure à filaments tégumentaires et à plumes ;
- et celui d'un oiseau ancien, Confuciusornis.
Toute présentation classique de la paléontologie et de la taphonomie (les processus de la fossilisation) souligne en général le caractère exceptionnel de la préservation des tissus mous et des molécules organiques originelles des organismes. Les exemples les plus classiques en sont les mammouths congelés dans les sols sibériens et quelques lambeaux de peau momifiés dans des bitumes naturels... et les plumes, tissus semi-durs, kératinisés, des dinosaures et des oiseaux.
Mais ces tissus ne conservent pas, en général, leur pigmentation. Comment cette coopération entre chercheurs de l'Académie des Sciences chinoise et de l'université de Bristol a-t-elle pu accéder à la couleur de ces fossiles ?
Leur démarche initiale a consisté à comparer les "filaments tégumentaires" de Sinosauropteryx et Sinornithosaurus et les plumes de Confuciusornis à l'échelle microscopique, par des observations au microscope électronique à balayage (MEB). Ils ont ainsi découvert, dans les plumes comme dans les filaments, de petites structures rondes, oblongues ou en ellipsoïdes très allongés, de taille sub-cellulaire.
Ces formes évoquent évidemment des bactéries, mais peuvent correspondre aussi à des dépôts minéraux apparus au cours de l'enfouissement et de la consolidation du sédiment (diagenèse). Il peut s'agir enfin d'organites intracellulaires conservés et c'est à cette dernière conclusion que sont parvenus les chercheurs. Ces structures seraient des mélanosomes, les vésicules qui stockent les deux principaux pigments cutanés (produits par des cellules dermiques spécialisées, les mélanocytes, à partir de la tyrosine) : l'eumélanine, noire, et la phæomélanine, rouge-brune (cf. figures 4 et 5a).

Source - © 2009 D'après Palaeobiology and Biodiversity Research Group [10] et http://photoprotection.clinuvel.com/node/204, modifiés
Ce diagnostic se fonde d'une part sur des publications précédentes, dans des revues plus spécialisées, qui montrent que les mélanosomes sont effectivement des structures très résistantes aux dégradations physico-chimiques liées à l'enfouissement et aux processus diagénétiques, et d'autre part sur des comparaisons directes avec des plumes actuelles (figure 5).
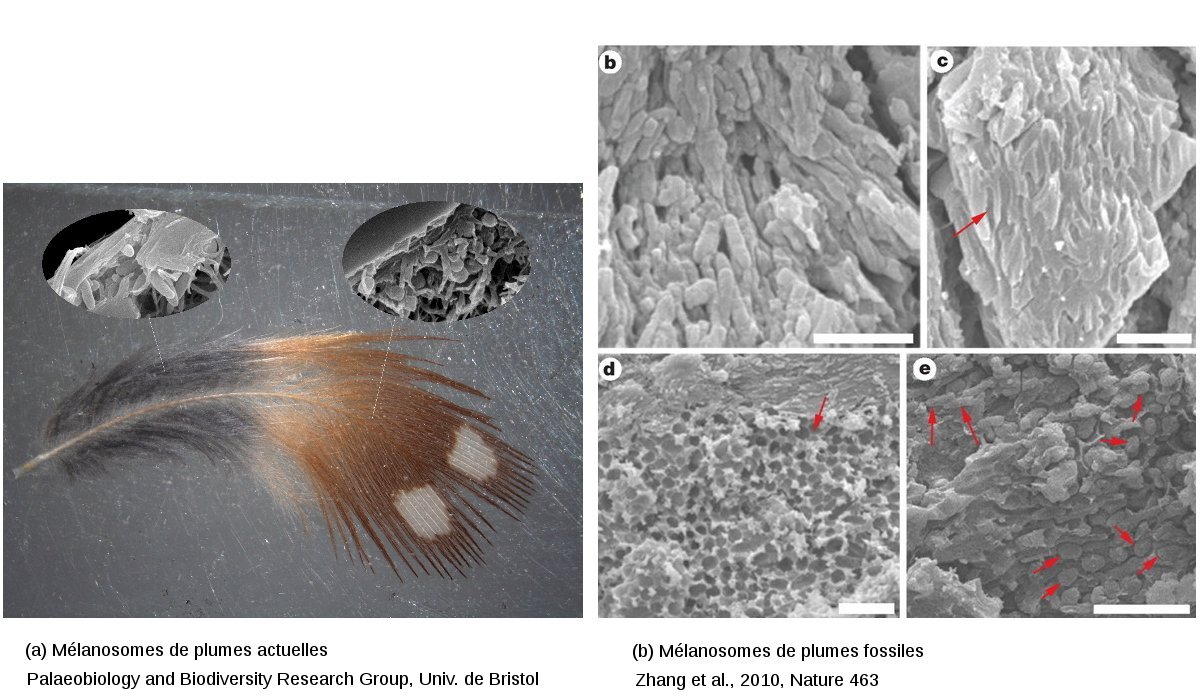
Figure 5. Mélanosomes de plumes actuelles et observations sur les fossiles
(a) Eumélanosomes (à eumélanine, dans les zones noires) et phæomélanosomes (à phæomélanine, dans les zones brunes) observées au MEB dans une plume de diamant mandarin de Timor (Taeniopygia guttata castanotis) actuel.
(b) Structures observées au MEB dans les plumes de Confuciusornis. Les flèches repèrent les corps alignés allongés, préservés à l'état de moulage et interprétés comme des eumélanosomes ou des phæomélanosomes [4]. Les barres d'échelle représentent 2 μm.
Sur l'image de droite, les clichés b et c sont interprétés comme des eumélanosomes bien alignés. L'image d correspondrait à des phæomélanosomes, moins bien ordonnés, surmontés d'une couches d'eumélanosomes. La photographie e montre le passage progressif d'une zone à eumélanosomes dominants (flèches longues) à une région de phæomélanosomes (flèches courtes).
Par ailleurs, les auteurs de l'étude avancent plusieurs arguments pour rejeter les deux autres possibilités, celles de structures bactériennes ou de minéraux, dont les suivants :
- ces corps allongés ne constituent pas une couverture superficielle, à l'instar d'un film bactérien, mais sont insérés à l'intérieur du corps de la plume, dans une matrice. Cette disposition, et les endroits de la plume (ou du filament), où ces corps sont accumulés, correspondent à ceux des mélanosomes des oiseaux actuels (qui s'organisent en une ou plusieurs couches au sein d'une matrice de kératine).
- la chimie des fossiles exclut l'origine minérale, en particulier la possibilité de pyrite en amas (pyrite framboïde), la taille des objets ne correspondant pas non plus à ce type de cristallisation.

Figure 6. Le théropode à « fourrure » de plumes, Sinosauropteryx
(a) Fossile de Sinosauropteryx conservé au Inner Mongolia Museum, Mongolie.
(b) Reconstitution du théropode Sinosauropteryx d'après les résultats de Zhang et al. [4].
Notons au passage que cette identification confirme que les filaments de Sinosauropteryx sont bien des plumes. Par ailleurs, les chercheurs ont pu ensuite examiner la répartition des deux types de mélanosomes dans ces organismes, puisque ceux-ci paraissent en place. Selon la quantité de mélanosomes présents et les proportions relatives des deux types, il devient possible d'établir une part de la coloration de l'animal, en tons de blanc (pas de mélanosomes), roux (phæomélanine dominante) et noir (eumélanine dominante). Ainsi, la queue de Sinosauropteryx ne contient que des phæomélanosomes et devait donc avoir une couleur « noisette à rouge » [4] (cf. figure 6b). Les plumes de Confuciusornis, quant à elles, présentent des changements rapides des proportions relatives des deux sortes de mélanosomes et devaient donc arborer des nuances de couleurs noires et brunes.
Évidemment, soulignent les auteurs, ils ne peuvent reconstruire mieux que « certains aspects de la coloration » : les oiseaux actuels possèdent souvent des couleurs beaucoup plus variées et d'innombrables nuances de teintes. D'autres pigments entrent en jeu, ainsi que la microstructure des tissus (la répartition plus ou moins ordonnées des fibres de collagène, par exemple), qui produit des « couleurs structurales » par diffraction différentielle des longueurs d'ondes lumineuses.
La parure d'Anchiornis huxleyi
Quelques semaines seulement après ces premiers résultats, une équipe sino-américaine remet en avant le Troodontidé Anchiornis huxleii en proposant une reconstitution complète de son plumage [5]. Cette étude se veut plus exhaustive et plus minutieuse que celles publiées dans Nature. Les auteurs ont d'abord effectué une analyse statistique (une analyse discriminante canonique) sur la forme, la taille, la densité de répartition et la distribution des différents mélanosomes dans les plumes d'un large échantillon d'oiseaux actuels, afin d'en tirer les paramètres les plus pertinents pour identifier précisément chaque catégorie de mélanosomes. Ils ont ensuite appliqué leurs résultats à plusieurs échantillons (29) prélevés en divers endroits sur le même fossile, celui d'Anchiornis.
Leurs conclusions sont que le corps de cet animal était à dominante noire et grise, mais arborait de larges zones blanches sur les longues plumes des membres antérieurs et postérieurs, ainsi qu'une houppe rouge associée à quelques plumes rouges sur les joues (cf. figure 7).
![Le nouvel aspect du Troodontidé Anchiornis, tel que proposé par Li et al. [5]](https://planet-terre.ens-lyon.fr/planetterre/objets/Images/plumes-dinosaures-oiseaux/couleurs-Anchiornis-huxleyi.jpg)
Source - © 2009 Michael DiGiorgio, Univ. Yale / National Geographic
Ainsi, selon les auteurs de ce dernier travail, les longues plumes des membres auraient eu très tôt, presque dès leur apparition chez les premiers Aviales, un aspect distinct de celles qui recouvraient le reste du corps. La surface supplémentaire automatiquement fournie par le développement de ces longues plumes aurait ainsi très vite été exploitée comme support à d'autres adaptations, peut-être lié à des processus de sélection sexuelle et à des comportements de type parade nuptiale. À moins que cette sélection sexuelle n'ait été la cause première du développement de ces longues plumes et que leur usage pour la course puis le vol, plané ou battu, n'en soit que des "produits dérivés" ?
Conclusion
La paléontologie des oiseaux et de leurs parenté dinosaurienne a ainsi connu un regain d'actualité, sur deux axes différents, représentatifs de deux aspects majeurs de la recherche paléontologique : 1) la détermination des phylogénies et des histoires évolutives d'une part, 2) la reconstitution des organismes eux-mêmes et de leurs particularités morphologiques et écologiques d'autre part. Ces deux pans de la recherche bénéficient tous deux des progrès des techniques : informatique (pour le calcul des arbres phylogénétiques) et imagerie. Mais comme toujours en paléontologie des vertébrés, les progrès restent dépendant de la découverte de nouveaux fossiles et de l'état de leur préservation : les progrès des connaissances depuis les années 1990 n'auraient jamais eu lieu sans la découverte des gisements exceptionnels de Chine.
On soulignera, pour finir, l'immense attrait médiatique que conservent encore et toujours dinosaures et oiseaux. Peu de recherches paléontologiques peuvent espérer susciter un jour une "course à la publication" comme celle qui vient d'opposer les deux revues scientifiques les plus en vue.
Références
- [1] Dongyu Hu, Lianhai Hou, Lijun Zhang, Xing Xu, 2009. A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature, 461, 640-643. doi:10.1038/nature08322
- [2] Lawrence M. Witmer, 2009. Feathered dinosaurs in a tangle. Nature, 461, 601-602. doi:10.1038/461601a
- [3] Jonah N. Choiniere, Xing Xu, James M. Clark, Catherine A. Forster, Yu Guo, Fenglu Han, 2010. A basal alvarezsauroid theropod from the early late Jurassic of Xinjiang, China. Science, 327, 5965, 571 - 574. doi:10.1126/science.1182143
- [4] Fucheng Zhang, Stuart L. Kearns, Patrick J. Orr, Michael J. Benton, Zhonghe Zhou, Diane Johnson, Xing Xu, Xiaolin Wang, 2010. Fossilized melanosomes and the colour of cretaceous dinosaurs and birds. Nature, 463,1075-1078. doi:10.1038/nature08740
- [5] Quanguo Li, Ke-Qin Gao, Jakob Vinther, Matthew D. Shawkey, Julia A. Clarke, Liliana D'Alba, Qingjin Meng, Derek E. G. Briggs, Richard O. PrumLi, 2010. Plumage color patterns of an extinct dinosaur, Science, 327, 5971, 1369 - 1372. doi:10.1126/science.1186290
- [6] Kevin Padian, Armand de Ricqlès, 2009. L'origine et l'évolution des oiseaux : 35 années de progrès. Comptes Rendus Palevol, 8, 2-3, 257-280. doi:10.1016/j.crpv.2008.11.007
- [7] Ronan Allain, Émilie Läng, 2009. Origine et évolution des saurischiens. Comptes Rendus Palevol, 8, 2-3, 243-256 doi:10.1016/j.crpv.2008.09.013
- [8] Sankar Chatterjee, R. Jack Templin, 2007. Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. PNAS, 104, 5, 1576-1580. doi:10.1073/pnas.0609975104
- [9] Xing Xu, Fucheng Zhang, 2005. A new maniraptoran dinosaur from China with long feathers on the metatarsus. Naturwissenschaften, 92, 4, 173-177. doi:10.1007/s00114-004-0604-y
- [10] Pages du Palaeobiology and Biodiversity Research Group sur les mélanosomes, Université de Bristol, U.K.
Voir aussi
- Xing Xu, Xiaoting Zheng, Hailu You, 2009. A new feather type in a nonavian theropod and the early evolution of feathers. PNAS, 106, 3, 832-834. doi:10.1073/pnas.0810055106
- Richard Prum, Alan Brush, 2005. Les plumes des dinosaures. dossier Pour la Science n°48, Le monde des dinosaures

