Article | 30/11/2011
Australopithecus sediba, nouvel australopithèque d'Afrique du Sud
30/11/2011
Résumé
En avril 2010, le magazine Science publiait la description d'un nouvel homininé, daté de moins de 2 Ma et attribué à une nouvelle espèce d'australopithèque, Australopithecus sediba, « descendant probablement d'Australopithecus africanus... [mais] partageant plus de caractères dérivés avec les premiers Homo qu'aucun autre australopithèque » [1]. Dix-sept mois plus tard, cinq nouveaux articles réunis dans un seul numéro de la même revue sont venus compléter les premières conclusions : ils présentent le contexte géologique, une datation améliorée et l'analyse détaillée de fossiles surprenant par leur combinaison de caractères considérés comme archaïques (typiquement autralopithèques) avec des traits dérivés, semblables à ceux du genre Homo. Un nouvel exemple de fossile « mosaïque », une situation déjà rencontrée chez d'autres groupes et à d'autres époques où se diversifient des taxons nouveaux (Tiktaalik lors de la « sortie des eaux » des Tétrapodes, ou les divers dinosaures à plumes, p. ex.).
Table des matières
Remarque. Les noms d'espèces, écrits dans la classification linéenne en italiques, à l'aide du nom de genre (pourvu d'une majuscule) suivi du nom spécifique (sans majuscule) — Genre espèce — sont généralement abrégés par la majuscule du genre et le nom d'espèce (G. espèce). Dans le cas des Homininés, la définition, dans les années 1990, du genre Ardipithecus amène les paléoanthropologues à distinguer les groupes Australopithèques et Ardipithèques par les abréviations Au. (p. ex. Au. sediba) et Ar. (Ar. ramidus) respectivement.
Introduction
C'est dans le numéro d'avril 2010 de la revue Science qu'une équipe internationale menée par des chercheurs de l'université du Witwatersrand (Afrique du Sud) a présenté les premières analyses de deux homininés fossiles découverts dans une grotte d'un massif dolomitique du Nord-Ouest de Johannesburg, à 15 km au NNE d'un autre site bien connu, Sterkfontein. Un premier article [1] décrivait les fossiles, tandis qu'un second se concentrait sur le contexte géologique du site [4].
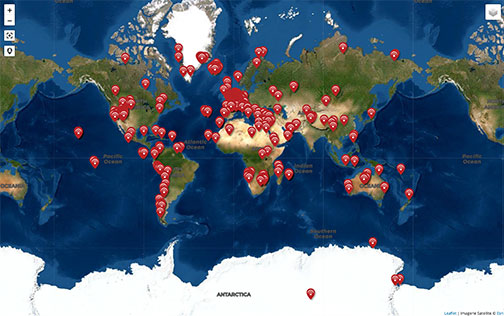
Ces deux nouveaux spécimens viennent compléter le riche registre d'Homininés extraits des formations sud-africaines, dans lequel figurait déjà plusieurs australopithèques sensu stricto, notamment Australopithecus africanus, et des australopithèques « robustes », ou Paranthropes (souvent considéré comme un groupe monophylétique issu des australopithèques), comme Paranthropus robustus (Figure 1).
Ces premières publications soulignaient déjà quelques originalités de ces nouveaux venus par rapport aux autres australopithèques connus, originalités qui conduisaient les auteurs à en faire une espèce nouvelle, Australopithecus sediba (sediba signifiant fontaine ou source en sesotho, une langue bantoue de cette région d'Afrique du Sud). Ils leur attribuaient par ailleurs un âge inférieur à 2 Ma (entre 1,95 et 1,78 Ma), ce qui en fait les plus récents représentants des australopithèques (si l'on en exclut les paranthropes).
Source - © 2011 Cyril Langlois
Figure 1. Répartition cartographique des fossiles d'Homininés mis au jour en Afrique
Au. sediba est signalé par un cartouche jaune.
Inspiré et complété de A. Gibbons, Science, 295, 2002
Une seconde série de cinq articles, parus dans la revue américaine en septembre 2011, est venue préciser et compléter les premières conclusions après plus d'un an d'études minutieuses de ces restes, en particulier de certaines structures anatomiques très informatives et bien conservées : le poignet, la main, le pied, la cheville et le pelvis. L'emploi de techniques sophistiquées de tomographie a également permis aux chercheurs de reconstituer un moule interne virtuel de la cavité crânienne (l'endocaste) de l'un des fossiles. Toutes ces structures montrent, selon ces chercheurs, un assemblage inattendu de caractères « type australopithèques » et « type Homo ». Enfin, des datations radiochronométriques et des analyses paléomagnétiques effectuées sur les roches sous- et sus-jacentes aux fossiles sont venues préciser leur âge : 1,977 ± 0,002 Ma [7].
Le site et le cadre géologique et chronologique
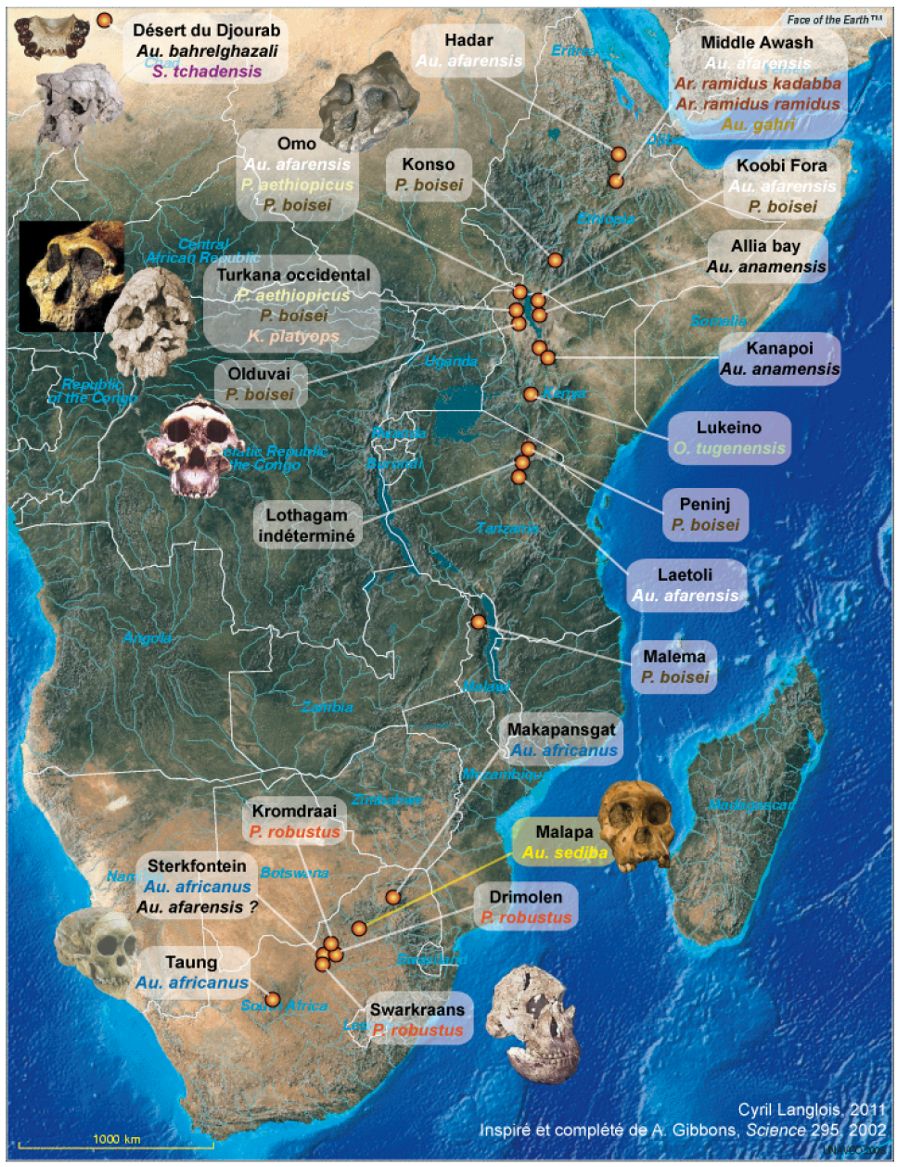
Les deux spécimens d'Au. sediba décrits à ce jour ont été extraits d'une cavité karstique remplie de sédiments fluviatiles, située dans des dolomies stromatolithiques datées de l'Archéen récent (2,64 à 2,5 Ga) et constituant la terminaison Nord-Est d'une série de cavités parallèle au litage de la dolomie, inclinée de quelques degrés vers le Nord-Nord-Ouest, mais d'un pendage légèrement inférieur à la pente du flanc de vallée sur lequel elle débouche. L'ensemble correspond à un ancien système karstique aujourd'hui érodé par la rivière Grootvleispruit (Figure 2).
Des sédiments clastiques, alternant avec des périodes d'érosion et des épisodes plus calmes marqués par la précipitation de calcaires, se sont accumulés dans la cavité et présentent cinq faciès distincts ; les fossiles ont été déterrés dans l'un d'eux, ainsi que des restes de chevaux, de félins, de hyènes, de lièvres et de micro-mammmifères. Les deux homininés paraissent avoir été apportés dans cette cavité, et ainsi mis à l'abri des charognards, par le même flux de sédiments, lesquels ont été rapidement lithifiés, d'où la bonne conservation des fossiles.
Le matériel fossilifère reposait sur des draperies calcaires (flowstone), présentant une aimantation rémanente inverse, alors que les sédiments porteurs des restes d'homininés montrent un polarité paléomagnétique normale ou intermédiaire. Les draperies ont pu être datées, à l'aide de la méthode U-Pb, par deux laboratoires distincts, situés l'un à Melbourne, l'autre à Berne, dont les résultats se sont révélés quasi-identiques, compte tenu des incertitudes de la méthode. Ils donnent un âge de 2,026 ± 0,021 Ma. Ces résultats suggéraient dès 2010 que les fossiles appartenaient à la biozone Olduvai, ce qui signifiait un âge compris entre 1,95 et 1,78 Ma [4]. Cette datation impliquait que ces australopithèques étaient possiblement contemporains des plus anciens représentants sud-africains du genre Homo, genre commençant, pour les auteurs des articles, avec Homo erectus. (Ces chercheurs suivent en cela l'avis des paléoanthropologues qui contestent l'appartenance des espèces H. habilis et H. rudolfensis au genre Homo et les considèrent comme des formes d'australopithèques [10]).
Source - © 2011 Dirks et al. / Pickering et al
Figure 2. Description du site de Malapa, Afrique du Sud
(a) Reconstitution du site de Malapa et du contexte de fossilisation des spécimens d'Au. sediba.
(b) Coupe NE-SW et ESE-OSO du site actuel. La ligne noire barbelée marque la base de la cavité dégagée selon l'axe de coupe.
D'après : (a) Dirks G. M. et al.[4], francisé ; (b) Pickering R. et al. [7], francisé.
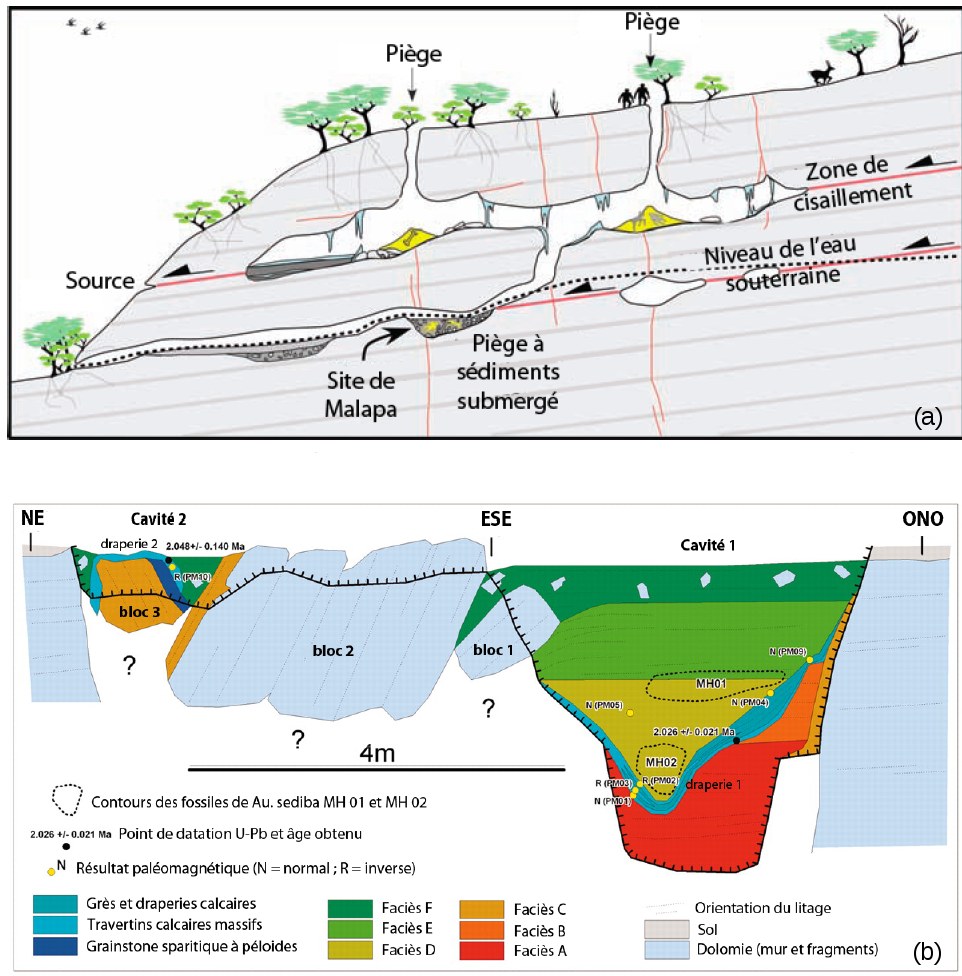
L'article de 2011 de R. Pickering et al. [7] reprend cette question de la datation d'Au. sediba. L'examen d'une nouvelle section du site (cavité 2 sur la figure 2b) a permis aux auteurs d'identifier dans les calcaires de brefs renversements du champ magnétique, d'une durée de quelques dizaines de milliers d'années ou moins (des excursions magnétiques) ; combinées à de nouvelles mesures isotopiques U/Pb, ces données les conduisent à placer la date de dépôt de ces australopithèques dans l'une de ces excursions et à restreindre ainsi l'âge de ces fossiles à 1,977 Ma.
Les fossiles sont en effet postérieurs à la draperie 1 de la cavité dans laquelle ils ont été trouvés, dépôt calcaire daté de 2 Ma et de polarité magnétique normale (Figure 2b) ; mais ils sont antérieurs à la draperie 2 de la seconde cavité, de polarité inverse. Par la méthode U/Pb, l'intervalle de dépôt entre les deux placages calcaires se trouve compris entre 1,91 et 2,05 Ma. La draperie 2 doit donc précéder l'épisode magnétique normal Olduvai, daté de 1,95 Ma. Enfin, le faciès D dans lequel ont été trouvés les fossiles montre une polarité intermédiaire. Or la seule fluctuation du champ magnétique connue avant l'évènement Olduvai est une excursion — une déviation marquée du champ magnétique, généralement accompagnée d'une chute de son intensité, de courte durée géologique mais n'aboutissant pas à une inversion complète — l'excursion pré-Olduvai [9], datée de 1,977 Ma (Figure 3). Au. sediba serait alors légèrement plus ancien que les premiers H. erectus. Pour les auteurs, cela implique que Au. sediba devient un ancêtre possible des premiers Homo.
Remarque. En théorie au moins, cette filiation restait envisageable même si les deux espèces avaient été contemporaines : dans un schéma d'évolution de type « équilibres ponctués », l'espèce (morphologique) ancestrale peut perdurer après l'apparition de sa (ou ses) espèce(s)-fille(s). Mais l'ancienneté plus importante de Au. sediba assure que cette espèce a effectivement pu être la forme ancestrale.
Source - © 2008 Roberts et al.
Cartouches noirs (resp. blancs) : période normale (resp. inverse). Flèches noires : inversions ; bleues : excursions reconnues ; rouges : excursions possibles, non validées (en 2008). L'excursion pré-Olduvai est soulignée.
D'après : Roberts A. P., 2008 [9], modifié.
Les fossiles
Le site fouillé a livré plusieurs homininés, mais deux seulement ont été dégagés et décrits pour l'instant. Les os exhumés sont attribués à un jeune mâle (MH1, défini comme l'holotype, c.-à-d. le spécimen de référence) et à une femelle adulte (MH2, moins complet et qui par conséquent devient un paratype, c.-à-d. un spécimen qui vient compléter la définition de l'espèce).
Selon Lee R. Berger et al. [1], plusieurs caractères anatomiques, comme par exemple les attaches musculaires bien marquées, attestent que MH1 était un mâle. En se basant sur les modèles de croissance de l'homme actuel et des grands singes, ces auteurs estiment que MH1 était « à 95% adulte », car ses deuxièmes molaires étaient déjà sorties et en contact avec leurs vis-à-vis (en occlusion). Dans un article ultérieur [2], les chercheurs estiment que « MH1 avait atteint un stade de développement équivalent à celui d'un enfant humain de 12 à 13 ans ». Cette conclusion est importante, car elle leur permet de supposer que même si MH1 n'avait pas achevé sa croissance, la poursuite de celle-ci n'aurait plus guère modifié les caractères morpho-anatomiques décrits sur le fossile ; autrement dit, les caractéristiques de MH1 seraient valables pour reconstituer l'anatomie adulte du mâle de cette espèce.
L'attribution de MH2 à une femelle est suggérée par la taille inférieure du squelette et de la mandibule de MH2 comparés à MH1 ; l'occlusion plus complète et l'usure plus prononcée des dents indiqueraient, de leur côté, un âge plus important et donc un adulte.
Ces deux individus montrent plusieurs traits dérivés par rapport aux espèces d'australopithèques antérieures, comme Au. anamensis ou Au. africanus. Par ailleurs, ils ne présentent pas les caractéristiques originales des paranthropes (ou australopithèques robustes : P. aethiopicus, P. robustus et P. boisei), comme la grande taille des prémolaires et des molaires, le développement des arcades zygomatiques, la face très large, alors que P. robustus et P. boisei sont datés de la même époque (entre 2 et 1 Ma environ) et que les spécimens de P. robustus viennent également d'Afrique du Sud. Au contraire, Au. sediba se signale par une petite taille absolue des prémolaires et molaires, qui rappelle plutôt les dents du genre Homo.
Plus de quatre pages de l'article de Berger et al. [1] sont occupées par une comparaison des caractères identifiés et des mesures effectuées sur Au. sediba avec ceux des autres australopithèques, des paranthropes et des anciens Homo, venant appuyer la conclusion des auteurs que ces restes sont ceux d'une espèce inédite. Plusieurs caractère rapprochent Au. sediba des espèces du genre Homo ; cependant, l'article de 2010 ne les considèrent pas suffisants pour placer cette espèce dans le genre Homo, comparés aux caractéristiques « australopithèques », notamment la petite taille corporelle (1,20 m environ, comme indiquée par la figure 4), le petit volume cérébral, les longs membres antérieurs, la forme des cuspides dentaires. Toutefois, aucune analyse phylogénétique n'est proposée dans cet article. Ces fossiles sont donc maintenu au « grade » australopithèque et ce groupe des australopithèques devient ainsi un peu plus un assemblage paraphylétique où sont regroupées les formes « non-Homo » ou « ni-Homo, ni-paranthropes ».
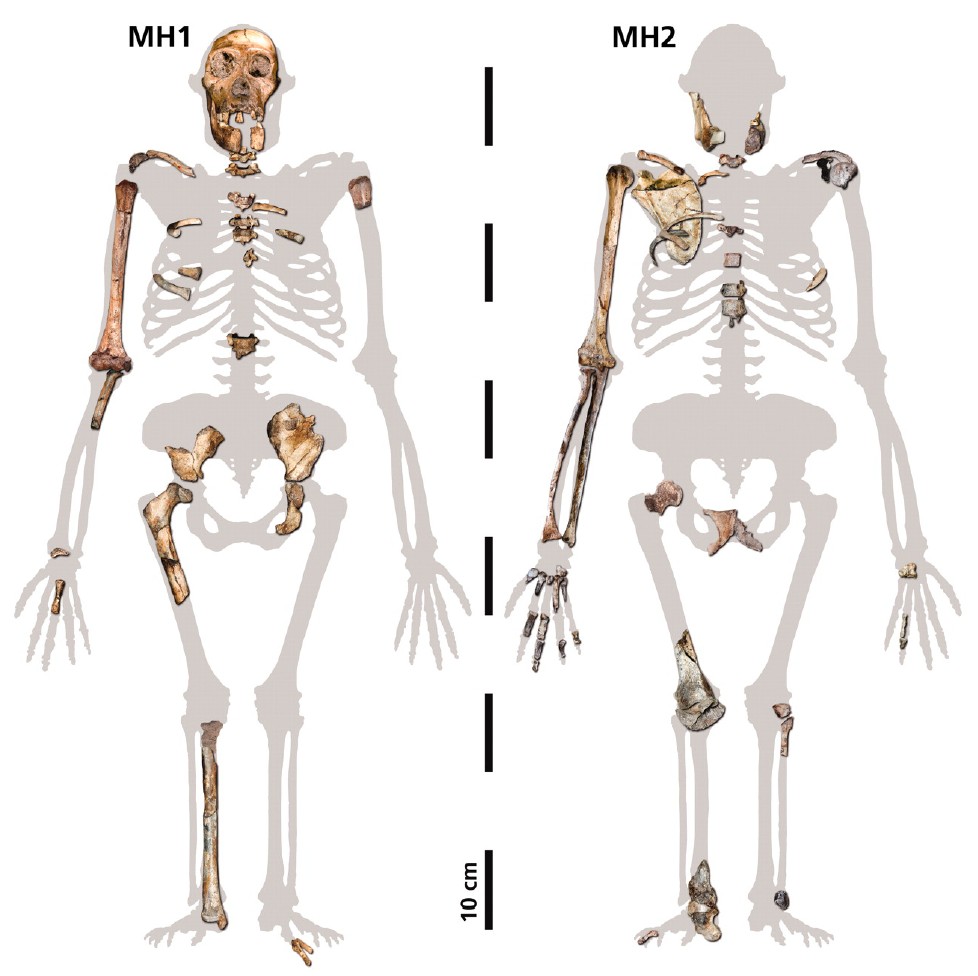
Source - © 2010 Berger et al.
Figure 4. Pièces exhumées des spécimens MH1 et MH2 d'Au. sediba
Les pièces de squlette ont été replacées à leur position anatomique approximative sur une silhouette d'Australopithecus africanus ajustée pour tenir compte des différences de proportions corporelles entre les deux espèces.
D'après : Berger L.R. et al., 2010 [1]
La figure 4 rassemble les fragments de squelettes dégagés : ces éléments sont particulièrement intéressants, puisqu'ils comprennent des os porteurs de nombreux caractères informatifs, par exemple plusieurs articulations (hanche, ceinture scapulaire, genou, cheville), un crâne et des portions de mandibule.
Quatre des cinq articles publiés en septembre 2011 sont ainsi consacrés à la description d'une de ces portions : pied et cheville [11], pelvis [5], main [6] et endocaste [2].
La main singulière d'Au. sediba
Les chercheurs disposent ainsi d'une main (droite) et du poignet correspondant, quasi-complets, de MH2 (Figure 5c) ; à cette structure s'ajoutent l'ensemble du bras droit et plusieurs os de la main gauche (Figure 4). La plupart de ces os sont complets et non déformés, et quelques autres os carpiens, ayant pu appartenir à MH1, viennent compléter cet ensemble exceptionnel. Ces derniers fragments permettent d'appréhender l'expression du dimorphisme sexuel sur cette structure rarement fossilisée : les métacarpes de MH1 s'avèrent ainsi plus robustes que leurs équivalents chez MH2.
La main d'Au. sediba présente une association originale d'états de caractères : des phalanges courbes comme chez les autres australopithèques, mais particulièrement longues ; un pouce de très grande taille par rapport aux autres doigts, sans doute bien musclé, à l'extrémité aplatie et muni d'une insertion tendineuse du muscle « long fléchisseur du pouce » bien marquée (Figure 5c), caractères qui ont été associés, chez d'autres fossiles, à la production d'outils. Prudents, Tracy Kivell et al. indiquent que cette main et ce pouce devaient pouvoir saisir des objets avec précision et les serrer fortement. Mais ils soulignent également que certains aspects de cette morphologie au moins ont été décrits chez des organismes plus anciens, comme Orrorin tugenensis (6 Ma) et Ardipithecus ramidus (4,4 Ma). « L'expansion des extrémités des doigts et l'insertion bien développée du muscle fléchisseur du pouce » seraient donc, écrivent-ils [6], « soit un état ancestral chez les homininés soit un convergence développée chez plusieurs taxons » (en relation ou non avec la fabrication d'outils), soulignant aussi que « ces caractères sont absents du pouce de OH7 » (code d'un fossile attribué à Homo habilis). Cependant, ajoutent ces auteurs, d'autres indices suggèrent que certains muscles de la main étaient moins développés chez MH2 que chez les homininés utilisateurs et fabricants d'outils et que, par ailleurs, main et poignet devaient permettre le grimper et la progression arboricole, quoique probablement moins efficacement que chez des homininés antérieurs (Au. afarensis), pour qui l'efficacité de la main dans les activités arboricoles étaient déjà restreinte par rapport à l'ancêtre commun des Hominidés.

Source - © 2011 Kivell T.L. et al., , modifié
(a) Structure du poignet chez l'humain moderne, en vue palmaire.
(b) Forme des phalanges en vue latérale chez les grands singes, les australopithèques et l'Homme actuel. (Benton, Vertebrate Palæontology, 2e éd., Blackwell Science, 2000).
(c) Main et poignet de l'individu MH2 d'Au. sediba, en vue palmaire (à gauche) et dorsale (à droite). Les états de caractères comparables à ceux rencontrés chez les Homo, donc dérivés, sont signalés en blanc, ceux de type australopithèque en jaune.
D'après Kivell T.L. et al. 2011 [6], modifié
T. Kivell et al. reconnaissent qu'il est particulièrement difficile de départager, sur les os, les caractéristiques morphologiques attribuables au mode de locomotion de celles liées à la fabrication d'outils et que les états de caractères liés à l'arboricolie n'interdisent pas la production d'outils. Ils soulignent cependant qu'Au. sediba possède des états de caractères jusque là connus seulement chez des espèces plus récentes, manufacturières d'outils ou supposées telle. Ainsi, une autre main fossile, le spécimen OH7, a été attribuée à H. habilis car retrouvée en association avec des outils de type oldowayien et donc supposée appartenir à l'auteur de ces objets ; cette conclusion est assez généralement acceptée, pourtant Au. sediba, plus âgé, montre une morphologie encore plus dérivée : il pourrait donc lui aussi être producteur d'outils. À l'inverse, expliquent-il, la morphologie du poignet de l'Homo floresiensis rappelle celle du chimpanzé, pourtant cet Homininé beaucoup plus récent qu'Au. sediba est clairement associé à une industrie lithique. Ainsi, par sa mosaïque de caractères, Au. sediba obscurcit les liens supposés entre morphologie et comportements.
Pied et hanche : encore des mosaïques de caractères
Cette coexistence de caractères dérivés et ancestraux ne concerne pas que la main d'Au. sediba. L'ensemble cheville-pied-extrémité du tibia, également attribué à MH2 (Figure 4), montre lui aussi une structure inédite. C'est par ailleurs la première fois que ces os sont retrouvés en connexion chez un homininé fossile. Pour ne pas les séparer, et éviter ainsi de possibles détériorations, les chercheurs ont reconstitué la structure, emprisonnée dans sa matrice sédimentaire, par un dispositif de tomographie médicale.
Il apparaît ainsi que l'articulation de la cheville de MH2 (la jonction talon-os crural) rappelle celle de l'Homme par la distribution des os autour de la surface d'articulation et signale l'adaptation à la bipédie : le tibia est perpendiculaire au grand axe du pied, et le genou, par conséquent, au-dessus de ce dernier (et non en avant de lui). De même, Au. sediba aurait pu avoir un pied arqué, quoique les indices en ce sens soient minces, et une insertion des muscles du mollet (le triceps sural) semblable à notre tendon d'Achille. Mais, en même temps, les analyses morphométriques détaillées rapproche le calcaneus d'Au. sediba de celui des chimpanzés plutôt que de ceux de l'Homme. De même, les auteurs [11] s'étonnent de la position d'une protubérance du calcaneum. Celle-ci évoque plus l'anatomie du Chimpanzé que celle de l'Homme ou même d'un autre australopithèque plus ancien, Au. afarensis. Se pose ainsi le problème de l'évolution plus ou moins complexe du pied, et de possibles convergences : si Au. sediba est plus proche d'Homo que Au. afarensis, il y a convergence, pour ce caractère, entre ces deux derniers. Si Au. sediba est plus éloigné d'Homo qu'Au. afarensis, les autres caractères dérivés du pied de Au. sediba seraient des convergences avec le genre Homo. Encore une fois, les auteurs ne proposent pas d'analyse phylogénétique pouvant tester ces scénarios alternatifs.
Quoi qu'il en soit, le pied d'Au. sediba ne devait pas pouvoir supporter le poids du corps aussi efficacement que celui de l'Homme moderne. Sa bipédie s'accompagnait alors vraisemblablement d'adaptations posturales, dynamiques, difficiles à concevoir à partir de ces seuls restes. Et quelle que soit la position phylogénétique exacte de ce nouveau fossile, l'évolution du pied et de la locomotion chez les Homininés a manifestement suivi un schéma complexe.
Le constat est quasiment le même concernant le pelvis, que Kibii et al.[5] ont pu reconstituer partiellement sur les deux individus. Comme dans le cas de la main, l'une des premières difficultés est de trouver des fossiles à comparer à ceux-ci et d'âge relativement voisin : les auteurs ont pu se baser sur un pelvis éthiopien de moins de 1,4 Ma — mais sans restes crâniens associés, donc difficile à attribuer à un Homo plutôt qu'à un Australopithecus — ainsi que sur les pièces de quelques Homo du Pleistocène ancien, comme l'adolescent du Turkana (KNM-WT 15000).
Le bassin d'Au. sediba partage lui aussi plusieurs caractères apomorphes —ou convergents— avec ceux du genre Homo, tout en conservant des caractéristiques australopithèques (Figure 6). D'autres caractères paraissent « intermédiaires » entre les deux pôles humain et australopithèque. Ainsi, l'os de la hanche aurait de nombreux caractères plésiomorphes (australopithèques). Les auteurs suggèrent aussi que « cette espèce ne présentait pas encore le retard de la fusion des apophyses pubiennes qui contribuent, chez l'Homme actuel, à accentuer le dimorphisme sexuel du bassin ». Au niveau du bassin, les Au. sediba présenteraient moins de différences entre les deux sexes que l'Homo sapiens 1.
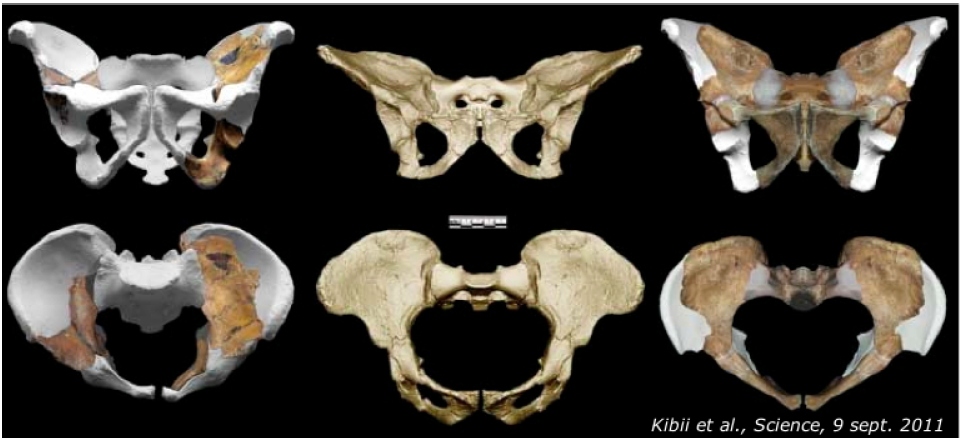
Source - © 2011 Kibii et al.
Job Kibii et ses collègues [5] discutent des caractéristiques de ces bassins dans la perspective du débat qui oppose deux hypothèses classiques avancées pour interpréter les transformations du bassin intervenues chez Homo et chez les australopithèques :
- la première hypothèse, « cinématique », considère que le bassin des australopithèques ne permettait qu'une bipédie assez proche de celle du chimpanzé (lorsqu'il se tient debout), parce que le mode de vie partiellement arboricole des australopithèques maintenait une sélection stabilisante en faveur des capacités à grimper : la bipédie australopithèque aurait été un compromis entre capacités arboricoles, qui imposent une grande souplesse au bassin, et marche terrestre ; chez Homo, les modifications du bassin auraient été permise par la restriction des modes locomoteurs à la marche terrestre et la spécialisation dans la bipédie au sol ;
- l'autre hypothèse, « obstétrique », suppose qu'une part au moins des transformations du bassin chez Homo est la conséquence de l'augmentation de la taille du crâne par rapport aux australopithèques et des contraintes consécutives sur la parturition.
La petite capacité crânienne d'Au. sediba était faible et rien ne permet de supposer que les nouveaux-nés de cette espèce avait un crâne plus gros à la naissance que les autres australopithèques. Dans le cadre de la seconde hypothèse, on pourrait alors supposer que son bassin montre les états de caractères plésiomorphes des australopithèques. Pour les auteurs, donc, « des traits dérivés de type Homo sur le pelvis peuvent apparaître indépendamment de l'encéphalisation et de l'accouchement de nouveaux-nés à gros cerveau ». Mais par ailleurs, l'importance fonctionnelle de ces traits dérivés de type humain n'est pas facile à estimer : autrement dit, si Au. sediba ne cadre pas avec le second scénario évolutif, il ne confirme pas clairement pour autant le premier. Malgré cela, Kibii et al. proposent de lier les transformations du bassin constatées chez Au. sediba par rapport à celui des australopithèques précédents à un changement des contraintes mécaniques, donc de la locomotion, elle-même probablement favorisée par une modification du mode de vie, « peut-être associée à l'expansion des prairies, au Plio-pléistocène, en Afrique du Sud ».
1 Certains médias ont écrit que le sexe femelle attribué à l'australopithèque Lucy avait été contesté et qu'il s'agirait d'un mâle. Comme expliqué dans l'ouvrage dirigé par Y. Coppens et P. Picq ([8], p. 257), une équipe suisse a seulement montré que, vu la petite taille du crâne du jeune australopithèque, la forme du bassin de Lucy ne traduisait pas nécessairement une adaptation liée à la parturition. Donc l'anatomie du bassin australopithèque n'est pas aussi informatif sur le sexe de l'individu que celui de l'homme moderne. Pour autant, cela ne suffit pas à récuser le statut femelle de Lucy.
L'endocaste d'Au. sediba
L'analyse des éléments du squelette post-crânien d'Au. sediba a fait largement usage des technologies récentes d'imagerie médicale et de reconstitution assistée par ordinateur. Il en est de même pour le crâne de MH1, dont l'intérieur a été partiellement recréé virtuellement à l'aide du rayonnement synchrotron (Figure 7b).
Le premier enseignement de cette étude est bien sûr le volume de ce moule cérébral : 420 cm3. Là encore, il est supposé que l'âge presque adulte de MH1 permet de considérer que la croissance de son cerveau était quasiment achevée. Ce volume endocrânien est du même ordre de grandeur que celui des autres australopithèques, et même plutôt dans la moitié inférieure de la gamme de variation des australopithèques. Évidemment, les auteurs considèrent cette petite taille comme une source d'étonnement compte tenu des états de caractères dérivés identifiés sur le squelette post-crânien et de la proximité qu'ils suggèrent avec le genre Homo. Toutefois, ils reconnaissent aussitôt après que ces « anomalies » pourraient s'expliquer par une réorganisation neurale indépendante d'un accroissement moyen de la taille du cerveau [2].
En dehors de son volume, c'est donc la forme de ce « moulage virtuel » de la cavité crânienne qui peut apporter des informations, même si, dans le cas présent, la majeure partie de l'hémisphère droit, des portions des lobes temporal et occipital gauches, et tout le cervelet, manquent. Qui plus est, les endocastes n'étant que des moules du cerveau, altérés encore par la fossilisation, il reste très difficile d'attribuer avec précision leurs bosses et leurs creux à des circonvolutions précises du cerveau. Il faut enfin noter que la forme du cerveau et l'évolution de cette forme sont liées aussi à l'évolution de la forme de l'ensemble du crâne, au travers du développement embryonnaire et post-embryonnaire : les transformations de la face ou de la base du crâne ont, elles aussi, des conséquences sur la forme du cerveau.
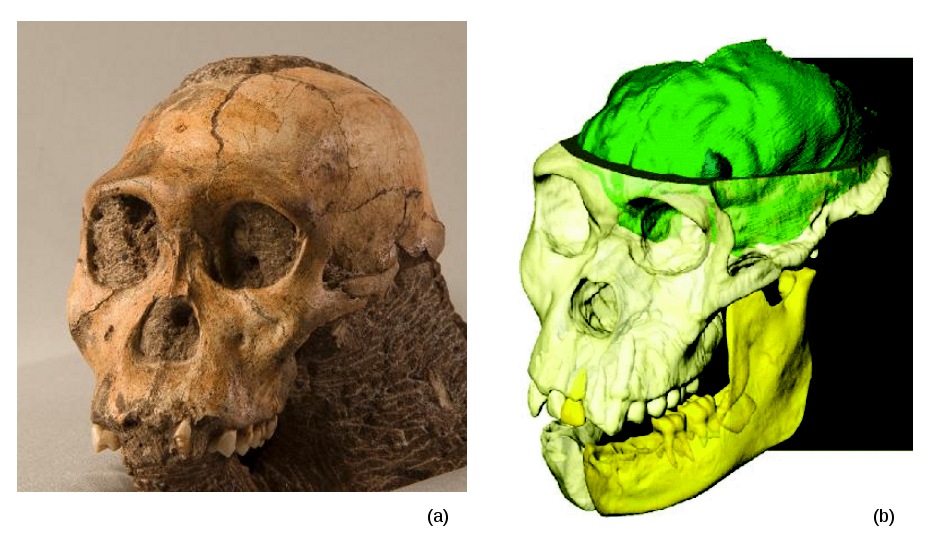
Source - © 2011 Berger - Eloff / Carlson et al.
Figure 7. Le crâne de l'Au. sediba MH1 et sa reconstitution numérique
(a) Crâne de MH1. Source : L.R. Berger, photographie : Brett Eloff
(b) Reconstitution virtuelle du crâne, de la mandibule et de l'endocaste de MH1. D'après : Carlson et al. , 2011 [2].
Cet endocaste partiel a néanmoins été comparé à ceux d'hommes actuels, de chimpanzés communs (Pan troglodytes) et d'australopithèques identifiés comme Au. africanus et Au. afarensis. Pour compléter leur échantillon, les auteurs ont considéré des hommes et des chimpanzés adultes, mais aussi des juvéniles présentant le même stade de développement dentaire que MH1. Une fois encore, ce nouvel Homininé montre des ressemblances nettes avec les australopithèques mais aussi quelques-unes avec l'Homme.
Pour quantifier un peu plus les comparaisons, des mesures morphométriques effectuées sur certains endocastes ont été reliées deux à deux sous forme de graphes bivariés et exploitées ensemble par analyse en composantes principales (ACP). Ces deux approches séparent clairement les chimpanzés de l'ensemble Homo - Australopithèques. Les graphes bivariés rapproche généralement Au. sediba de l'un des Au. africanus (Sts 5). L'analyse en composante principale sépare les hommes actuels des chimpanzés et rapproche les mêmes australopithèques Au. sediba et Sts 5 de l'humain (Figure 8).
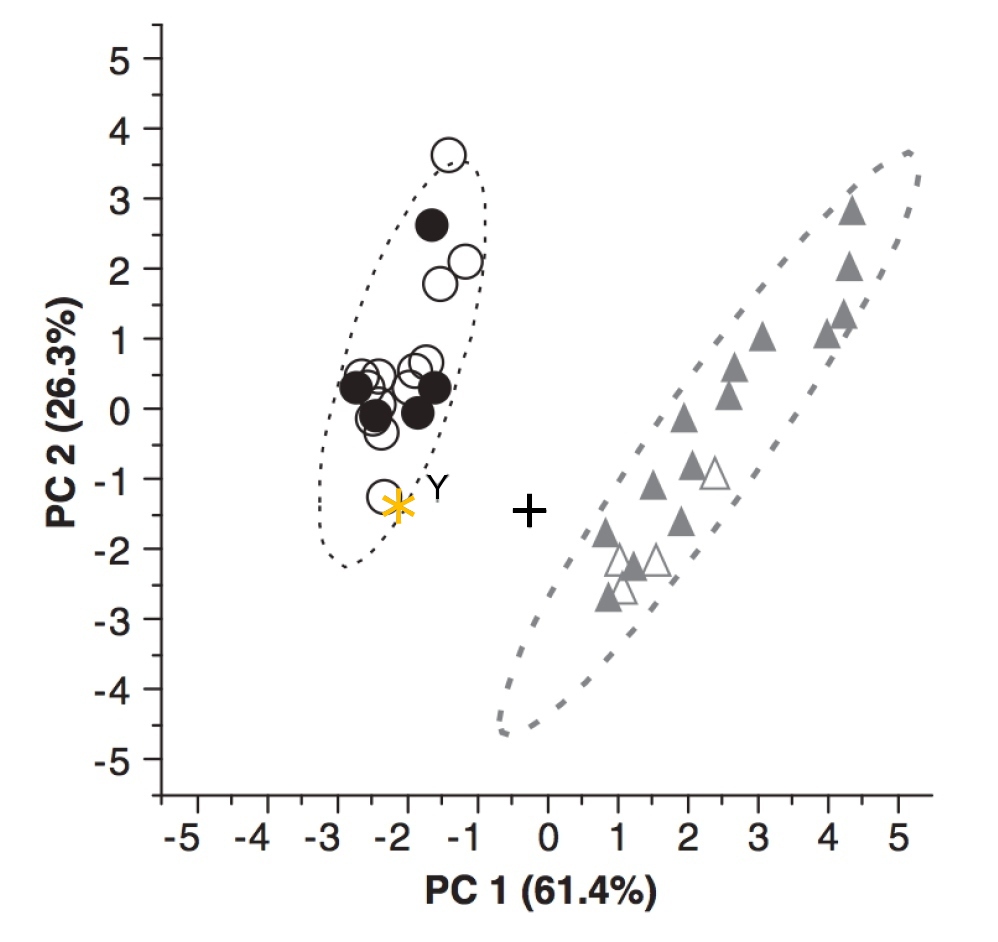
Source - © 2011 Carlson et al.
Les composantes, dans une ACP, ne correspondent pas à des mesures, mais à des combinaisons linéaires regroupant plusieurs mesures afin d'extraire les comlposantes principales, c'est-à-dire les paramètres qui permettent, ici, de distinguer Homo et chimpanzés. La position des australopithèques est donc vue dans un diagramme permettant de visualiser la proximité avec l'un ou l'autre de ces groupes.
La première composante principale PC1 explique 61,4% de la variance, la seconde, PC2, 26,2%. Chimpanzés et humains apparaissent bien séparé par PC1. L'alignement des points, visualisés par des ellipses de confiance à 95% nettement allongées, traduit une corrélation entre les deux composantes. MH1 entre — mais tout juste — dans l'ellipse de confiance des Homo sapiens actuels, contrairement aux deux autres australopithèques, plus anciens.
D'après : Carlson et al., 2011 [2].
Ainsi, Au. sediba se démarque peu des australopithèques au niveau de l'endocaste, même s'il montre quelques différences qui le rapproche de l'humain actuel. Il apparaît ainsi que ce nouvel homininé s'écarte beaucoup moins des australopithèques par sa morphologie crânienne que par celle du bassin ou de la main. On retrouve à nouveau ici le caractère mosaïque de ce fossile et la possibilité pour les diverses parties du squelette de changer à des vitesses différentes.
Conclusion
Comme dans d'autres cas, par exemple celui des tétrapodes primitifs et de leur « sortie des eaux », ce nouveau fossile démontre que la morphologie des organismes ne changent pas « d'un bloc » ni de façon homogène au cours du temps : les caractères considérés comme dérivés car réunis et bien marqués chez les organismes les plus récents peuvent n'être qu'en partie représentés chez les organismes précédents et y côtoyer des traits plus archaïques. Et rien ne permet de supposer que ces individus n'aient pas été tout aussi fonctionnels et adaptés à leur milieu que les précédents ou les suivants.
Ce cas-ci, ici encore plus marquant puisqu'il concerne « notre » branche évolutive, constitue donc un argumentaire concret contre l'argument classique des créationnistes pour qui un organisme « intermédiaire » ne peut exister car il ne serait pas viable. Australopithecus sediba, comme le proto-tétrapode Tiktaalik ou les dinosaures à plumes, démontre qu'il n'existe pas d'organisme « intermédiaire », mais des organismes mosaïques, chacun viable ; il souligne enfin que l'évolution morpho-anatomique est un processus complexe, où les différentes portions du squelette changent « séparément », à des degrés variables, et ce d'autant plus que tous ces changements n'ont pas nécessairement de conséquences fonctionnelles (donc ne sont pas des adaptations soumis à une sélection), mais peuvent résulter mécaniquement d'autres modifications (contraintes évolutives) ou n'être que des variations neutres.
Cet aspect disparate de l'évolution anatomique n'a pu être constatée ici que grâce à la préservation de plusieurs parties clés du squelette. Ce constat met donc aussi en garde contre les conclusions tirées de matériel par trop fragmentaire, pour lequel, comme le soulignaient Gilles Berillon et François Marchall dans un dossier Pour la Science de 2007, à propos de la bipédie [3], « la question [. . . ] est de savoir jusqu'à quel point un os seul peut nous permettre de déduire une fonction qui implique pourtant tout un système ».
Enfin, Australopithecus sediba souligne qu'évolution cognitive et évolution morphologique ne sont pas non plus forcément connectées ou proportionnelles : si la main d' Au. sediba autorisait des mouvements précis et la réalisation d'outil, son endocrâne n'en est pas plus large que ceux des autres australopithèques pour autant. On ne peut donc savoir si Au. sediba était effectivement concepteur et utilisateur régulier d'outils ; à l'inverse, cela relativise encore le lien classiquement réalisé entre outil et Homo (un lien déjà réfuté par Pascal Picq sur la base de l'observation des chimpanzés [3]). Si l'évolution du rameau humain en devient plus complexe à décrypter, elle n'en est que plus intéressante et se débarrasse, découverte après découverte, de sa vision anthropocentrée et linéaire et des conceptions simplistes auxquelles s'accrochent les mouvances créationnistes.
Références
- [1] Lee R. Berger, Darryl J. de Ruiter, Steven E. Churchill, Peter Schmid, Kristian J. Carlson, Paul H. G. M. Dirks, Job M. Kibii, 2010. Australopithecus sediba : A new species of Homo-like Australopith from South Africa, Science, 328(5975):195–204 - doi: 10.1126/science.1184944
- [2] Kristian J. Carlson, Dietrich Stout, Tea Jashashvili, Darryl J. de Ruiter, Paul Tafforeau, Keely Carlson, Lee R. Berger, 2011. The endocast of MH1, Australopithecus sediba, Science, 333(6048):1402–1407 - doi: 10.1126/science.1203922
- [3] Collectif, 2007. Sur les traces de nos ancêtres, Dossiers Pour la Science, n°57. Pour la Science
- [4] Paul H. G. M. Dirks, Job M. Kibii, Brian F. Kuhn, Christine Steininger, Steven E. Churchill, Jan D. Kramers, Robyn Pickering, Daniel L. Farber, Anne-Sophie Mériaux, Andy I. R. Herries, Geoffrey C. P. King, Lee R. Berger, 2010. Geological setting and age of Australopithecus sediba from Southern Africa, Science, 328(5975):205–208 - doi: 10.1126/science.1184950
- [5] Job M. Kibii, Steven E. Churchill, Peter Schmid, Kristian J. Carlson, Nichelle D. Reed, Darryl J. de Ruiter, Lee R. Berger, 2011. A partial pelvis of Australopithecus sediba, Science, 333(6048):1407–1411 - doi: 10.1126/science.1202521
- [6] Tracy L. Kivell, Job M. Kibii, Steven E. Churchill, Peter Schmid, Lee R. Berger, 2011. Australopithecus sediba hand demonstrates mosaic evolution of locomotor and manipulative abilities, Science, 333(6048):1411–1417 - doi: 10.1126/science.1202625
- [7] Robyn Pickering, Paul H. G. M. Dirks, Zubair Jinnah, Darryl J. de Ruiter, Steven E. Churchill, Andy I. R. Herries, Jon D. Woodhead, John C. Hellstrom, Lee R. Berger, 2011. Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo, Science, 333(6048):1421–1423 - doi: 10.1126/science.1203697
- [8] Pascal Picq, Yves Coppens, 2001. Aux origines de l'humanité. De l'apparition de la vie à l'homme moderne, volume 1. Fayard, Paris
- [9] Andrew P. Roberts, 2008. Geomagnetic excursions : Knowns and unknowns, Geophysical Research Letters, 35(17) - doi: 10.1029/2008GL034719
- [10] Bernard Wood, Mark Collard, 1999. The human genus. Science, 284:65–71
- [11] Bernhard Zipfel, Jeremy M. Desilva, Robert S. Kidd, Kristian J. Carlson, Steven E. Churchill, Lee R. Berger, 2011. The foot and ankle of Australopithecus sediba, Science, 333(6048):1417–1420 - doi: 10.1126/science.1202703