
Article | 16/05/2012
Le développement de la végétation continentale de l'Ordovicien au Dévonien et ses conséquences géologiques
16/05/2012
Résumé
Un récent numéro de la revue britannique Nature présente la ré-interprétation d'un site fossilifère dévonien à végétaux de l'état de New York (Gilboa). La présentation de ces nouvelles données fournit l'occasion de rappeler quelques étapes de la colonisation des terres émergées par les végétaux, entre l'Ordovicien et le Dévonien (gisement de Rhynie), et de présenter quelques idées et résultats récents sur les bouleversements occasionnés par cet épisode sur les enveloppes externes de la Terre.
Table des matières
Introduction
L'apparition des végétaux terrestres et leur expansion sur les terres émergées se sont déroulées de l'Ordovicien (488-443 Ma) au Dévonien (416-359 Ma). Les traces fossiles de ces événements sont nombreuses, et aux nouvelles découvertes viennent s'ajouter l'amélioration des connaissances ou les ré-interprétations de fossiles et de sites déjà connus. L'image de l'écosystème végétal qui occupait la région de Gilboa, dans l'état de New York, au cours du Dévonien supérieur, vient ainsi d'être renouvelée par l'étude de nouveaux affleurements mis au jour sur ce site découvert en 1870.
Parallèlement à ces progrès dans la reconstitution des communautés végétales, d'autres études se penchent sur les conséquences de cet épisode exceptionnel et irréversible. Nous verrons que les données de terrain, comme les modélisations analogiques et numériques, suggèrent que les petits végétaux multicellulaires puis le développement des formes arborescentes ont eu des effets multiples et profonds sur les enveloppes superficielles et qu'ils ont pu contribuer significativement à plusieurs événements géologiques encore mal compris, tels que la glaciation fini-ordovicienne ou l'extinction marine de la fin du Dévonien.
La colonisation végétale des terres émergées
Les traces fossiles de l'évolution des végétaux terrestres
Le registre fossile des premiers végétaux terrestres consiste largement en micro-fossiles et en débris macroscopiques, surtout pour les temps les plus anciens. L'une des difficultés est donc de les distinguer d'autres structures unicellulaires enkystées non végétales, ou des hyphes mycéliens, qui apparaissent à la même époque (figure 1).
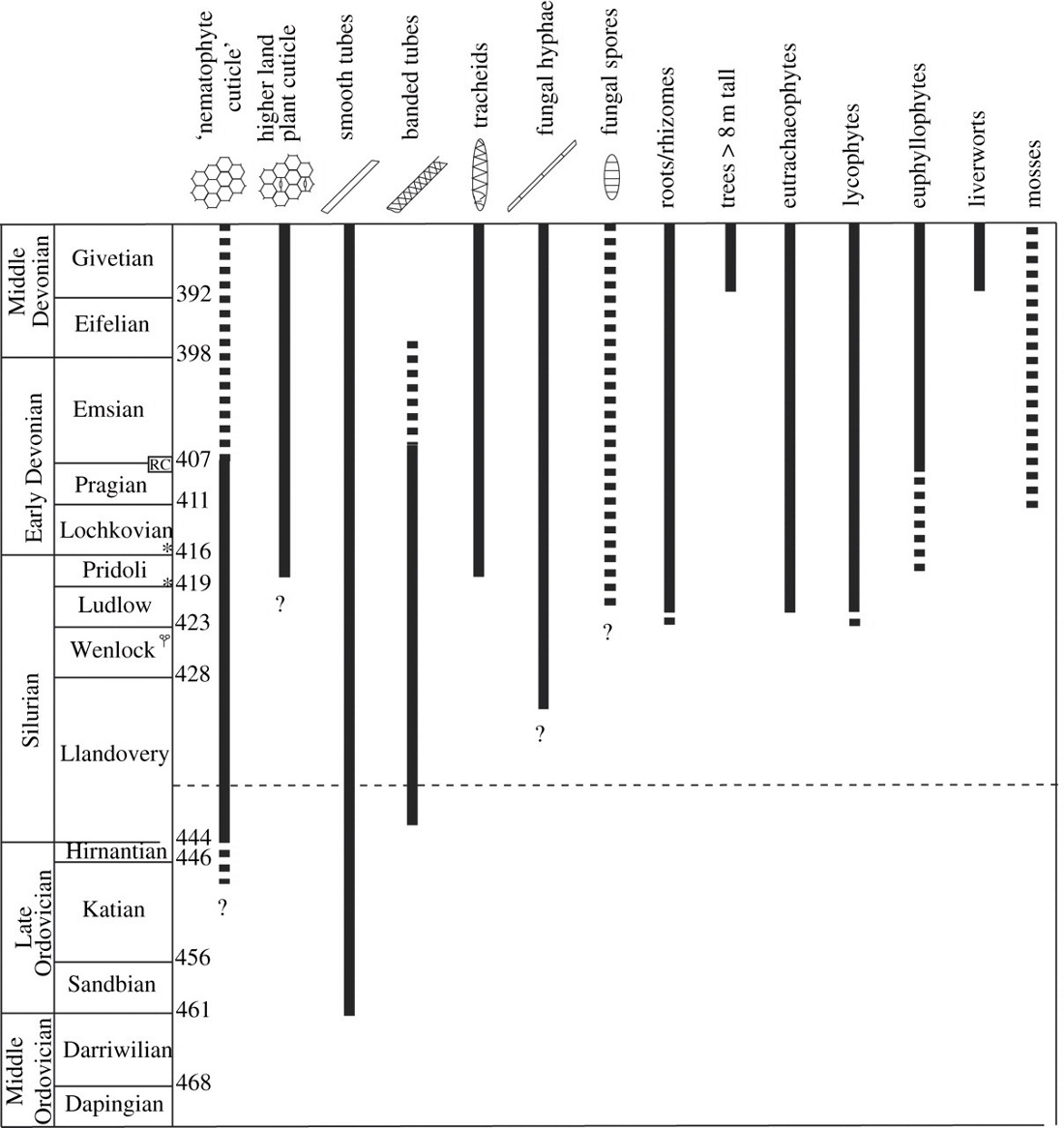
Source - © 2012 Kenrick et al. (Ref6)
Les barres pleines recouvrent les fossiles bien datés et déterminés, les barres en pointillés les fossiles d'identification douteuse. Les figurés dans l'échelle stratigraphique de gauche indique les horizons à fossiles végétaux majeurs. RC : cherts de Rhynie ; Lochkovien : assemblage carbonisé de North Brown Clee Hills ; Pridoli : assemblage carbonisé de Ludford Lane ; petit symbole “Cooksonia” au dessus de l'étage Wenlock : premiers restes de Cooksonia (Wenlock, Irlande).
Les micro-fossiles végétaux qui se rencontrent dès l'Ordovicien moyen sont des spores, qualifiées de cryptospores faute de pouvoir les attribuer à une espèce. Les plus anciennes ne montrent aucune structure de déhiscence et sont dites alètes. Elles se présentent sous forme de cellule seule (monade), ou d'association de deux (dyades) ou quatre (tétrades) cellules, nues ou enserrées dans une paroi commune, ce qui suggère un processus de méiose (figure 2). Certaines spores monocellulaires présentent un pourtour épais et plus clair : qualifiées de cryptospores hilates, elles sont interprétées comme des vues en coupe transversale d'une dyade (Gensel, Ref4). À partir de l'Ordovicien supérieur puis surtout au Silurien et au Dévonien se rencontrent des spores trilètes, à trois fentes de déhiscence.
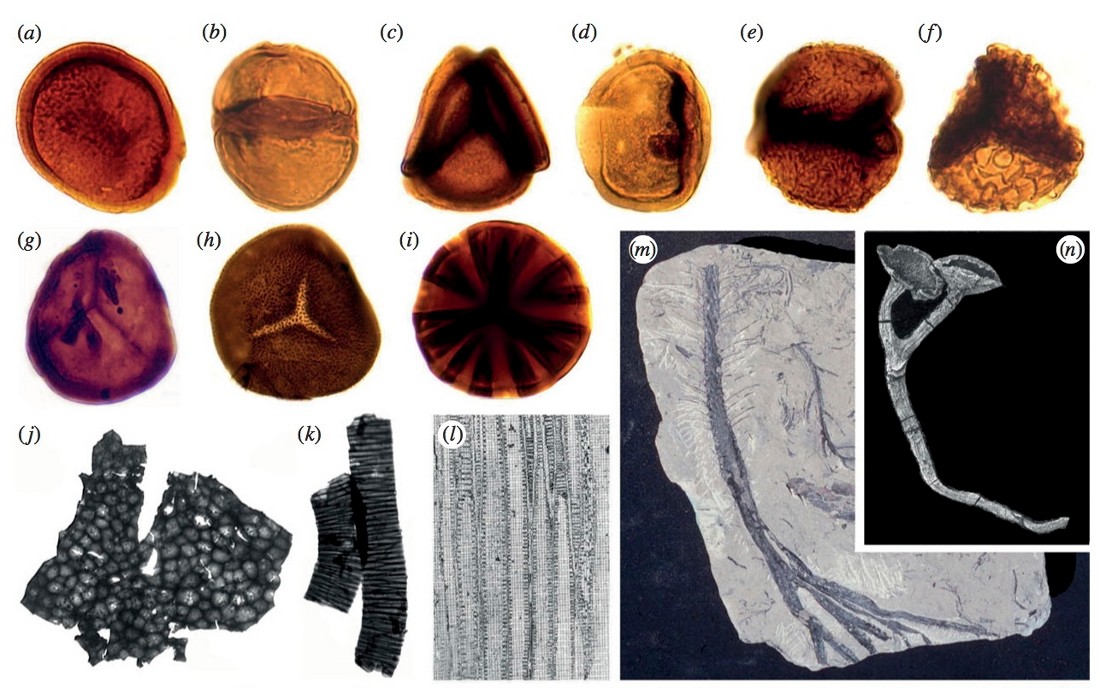
Source - © 2012 Kenrick et al. (Ref6)
Figure 2. Exemples de fossiles des premiers végétaux des terres émergées
(a) à (i) : cryptospores. (a) monade nue, (b) dyade nue, (c) tétrade nue, (d) monade enclose dans une enveloppe, (e) dyade à enveloppe, (f) tétrade à enveloppe, (g) spore trilète du Dévonien inférieur du bassin anglo-gallois. (h) à (i) : spores trilètes ornementées des sédiments du Dévonien inférieur associés aux cherts de Rhynie. (j) et (k) : débris végétaux dispersés du Dévonien inférieur de Rhynie. (l) tissus conducteurs de la plante vasculaire primitive Gosslingia (Dévonien inférieur). (m) système racinaire du Dévonien supérieur de Chine Chamaedendron multisporangium. (n) Cooksonia, Dévonien inférieur du bassin anglo-normand.
Au Silurien, les fossiles de plantes restent de petite taille ou parcellaires et relativement peu fréquents, avec moins d'une quarantaine de sites fossilifères. La première plante terrestre reconnue est la petite Cooksonia, découverte dans le Silurien d'Irlande (photo (n) de la figure 2).
C'est au Dévonien que le nombre de sites et la diversité des végétaux s'accroît nettement, avec, entre autre, l'emblématique site de Rhynie (Écosse), ou celui de Gilboa (état de New York, USA).
Le gisement de Rhynie
Le site de Rhynie est un gisement exceptionnel (un lagerstätte) situé au Nord-Est de l'Écosse, non loin d'Aberdeen, et daté du Dévonien inférieur. Il correspond à une cinquantaine de couches fossilifères, dont le contenu a été impeccablement préservé par silicification hydrothermale. Ces horizons siliceux[1]ont fourni divers arthropodes (acariens microscopiques du sol, arachnides, mais aussi les plus anciens insectes connus - Ref18), des restes d'hyphes mycéliens impliqués, pour certains dans des symbioses lichéniques ou mycorhiziennes (Selosse et Tacon - Ref13) et bien sûr les plus anciennes formes connues de végétaux vasculaires à ramifications aériennes, comme Aglaophyton major ou Rhynia gwynne-vaughanii.
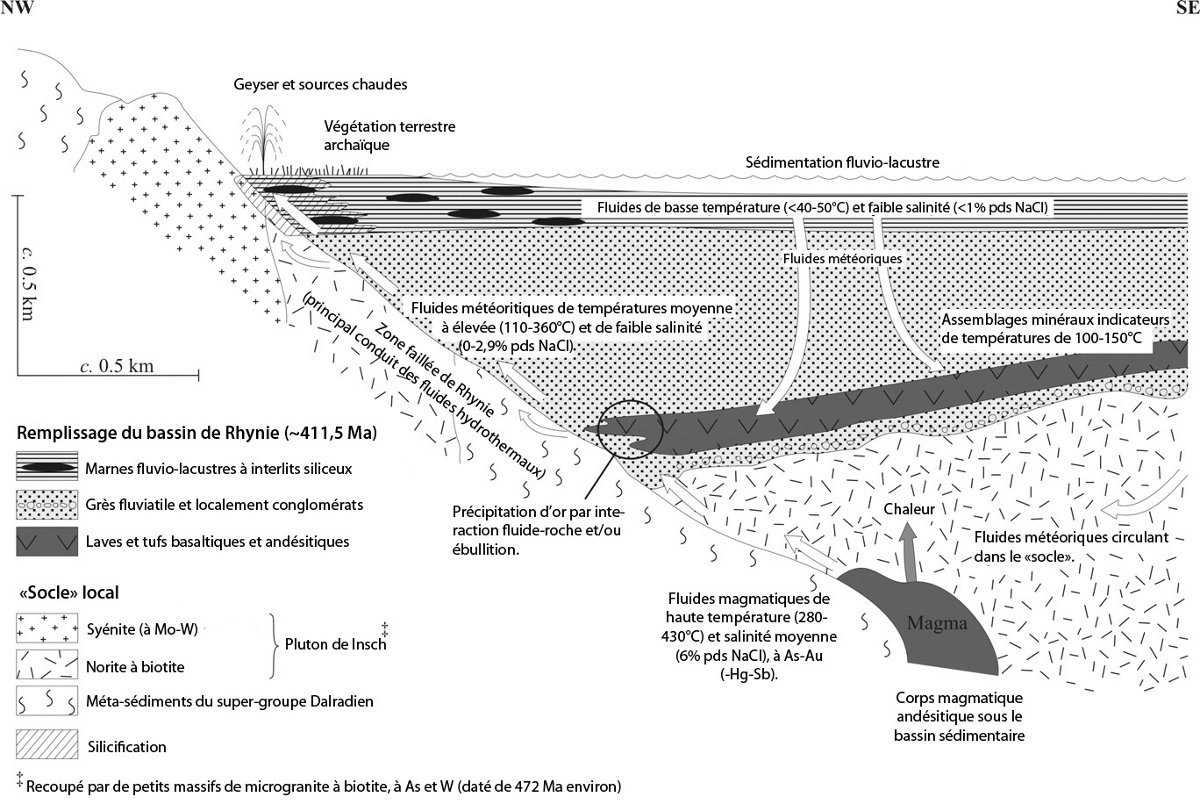
Il y a 411 Ma environ, Rhynie était une région marécageuse située en bordure d'un graben en extension associé à un volcanisme andésitique, où s'accumulaient, en contexte fluvio-lacustre, des sédiments détritiques (devenus aujourd'hui des grès et des schistes) et des produits volcaniques.
La circulation des fluides volcaniques et météoriques au sein de cet assemblage volcano-sédimentaire a solubilisé, concentré et re-déposé divers éléments minéraux économiquement intéressants provenant des sédiments volcaniques et détritiques — or (Au), étain (Sn), tungstène (W), etc — mais également la silice (figure 4). Ces fluides chargés de silice dissoute formaient des geysers en surface et leur refroidissement rapide provoquait la précipitation de la silice. Celle-ci a ainsi recouvert les organismes lacustres enfouis dans les couches marneuses, rempli les cavités cellulaires et imprégné les molécules organiques, peut-être en quelques années seulement, voire moins (Ref18). Ce type de précipitation siliceuse hydrothermale se rencontre aujourd'hui en Islande ou dans le parc de Yellowstone, par exemple, et ces dépôts siliceux sont justement dénommés geyserite. Par diagenèse, la silice initiale, micro-cristalline ou amorphe, dite opale-A, recristallise en opale-CT puis en quartz micro-cristallin (calcédoine).

Source - © 2011 Cyril Langlois
Figure 3. Cône de geyserite autour d'une source hydrothermale actuelle, Hveravellir, Islande
L'édifice au premier plan mesure environ 1,50 m de diamètre.
Les végétaux ainsi préservés à Rhynie sont de très petite taille, d'anatomie et de morphologie simples : des tiges dressées à ramification dichotomique, contenant un seul faisceau vasculaire central et portant des sporanges. En quelques dizaines de millions d'années, ces organismes vont se diversifier, et le site américain de Gilboa, du Dévonien moyen, donne un aperçu de cette évolution.
Source - © 2011 Parry et al., francisé (Ref12)
La végétation dévonienne de Gilboa : plus complexe qu'on ne croyait
C'est dans les années 1920, dans la carrière de Riverside (Riverside Quarry) et quelques sites voisins, près de la ville de Gilboa, dans l'état de New York, qu'ont été découverts de grands troncs transformés en moulages de grès. La formation gréseuse exploitée dans ces carrières est datée de -390 Ma, soit 21 millions d'années de moins "seulement" que Rhynie. Elle a pourtant livré la plus ancienne forêt fossile connue. L'espèce productrice des troncs fut baptisée Eospermatopteris. En 2007, d'autres fragments végétaux exhumés sur ce site (des tiges et des feuilles) furent identifiés comme des portions de la même plante (Ref15). Eospermatopteris était un végétal arborescent pouvant dépasser 6 m de hauteur, au tronc vertical, rectiligne, terminé par une couronne de feuille courtes. L'espèce est maintenant attribuée à la classe éteinte des Cladoxylopsides (Cladoxylopsida), proche des fougères.

Source - © 2012 Google earth
Figure 5. Positionnement géographique des sites fossilifères de Rhynie et de Gilboa
Sites fossilifères de Rhynie, en Écosse, et de Gilboa, état de New York, USA.
En 2010, l'évacuation des déblais de carrière a mis au jour une nouvelle portion de l'affleurement fossilifère de 1200 m2, correspondant à un paléosol. L'équipe de William E. Stein (Ref14) y a identifié les structures d'enracinement des Eospermatopteris, sous la forme de dépressions circulaires entourées d'un bourrelet de grès traversé par une couronne de racines rayonnant autour du creux central (figure 6).

Source - © 2012 Stein et al., in matériel supplémentaire (Ref15)
Le carroyage (fils tendus et touches de peinture orange) mesure 2,5 m (direction verticale) par 1,5 m (direction horizontale).
Mais en plus de ces systèmes racinaires, les auteurs de cette nouvelle étude ont identifié d'autres structures végétales, des tiges plus ou moins courbes, d'une quinzaine de centimètres de diamètres et de plusieurs mètres de longueur. Certaines montrent de fines excroissances latérales, interprétées comme des racines. Ces tiges se distinguent des Eospermatopteris par une couche externe carbonée de 1 à 3 mm d'épaisseur et la présence, bien reconnaissable au microscope, de faisceaux vasculaires à xylème primaire, xylème secondaire et phloème. Les chercheurs analysent ces restes comme les rhizomes d'un deuxième type végétal, qu'ils attribuent à l'ordre des Aneurophytales, lui-même inclus dans la classe des Progymnospermopsidés (Progymnospermopsida).
Enfin, parmi les fragments repérés sur cette dalle de grès figure encore une troisième espèce arborescente, rappelant certaines Lycophytes[2] connues dans des gisements plus récents, mais ici trop fragmentaire pour pouvoir être reconstituée ou attribuée à une espèce précise. La forêt de Gilboa comprenait donc au minimum trois espèces végétales de grande taille, appartenant à trois groupes taxinomiques différents.
Cette nouvelle fouille suggère également que ces différentes espèces, vu leur port et leur anatomie (tronc droit pour Eospermatopteris, rhizome serpentiforme pour les Aneurophytales) n'atteignaient pas les mêmes hauteurs et constituaient donc plusieurs strates arbustives, comme dans les forêts plus récentes : sous la canopée formée par les couronnes de feuilles des Eospermatopteris se développait un sous-bois d'Aneurophytales, dont les rhizomes au port lianescent se développaient horizontalement entre les arbres ou — comme le suggère la disposition de certains fossiles de Gilboa — montaient verticalement en prenant appui sur le tronc des Eospermatopteris. Les lycophytes arborescentes occupaient peut-être, quant à elles, un étage intermédiaire. L'organisation spatiale et écologique des forêts était donc, dès cette époque, tridimensionnelle.

Source - © 2012 Meyer-Berthaud et Decombeix (Ref10)
Figure 7. Reconstitution de la forêt de Gilboa
Cette image montre les grands Eospermatopteris arborescents, les rhizomes d'Aneurophytales constituant l'étage basal du sous-bois et une lycophyte occupant l'étage intermédiaire. L'apparence de la base et du sommet de cette dernière plante est inconnue. Voir aussi la vue d'artiste la forêt de Gilboa, en couverture du numéro de Nature du 1er mars 2012.
La végétation des terres émergées a donc considérablement évolué entre l'Ordovicien et le Dévonien moyen, et particulièrement au Dévonien, passant de petites plantes à de véritables forêts. Or on sait que les végétaux ont des effets importants sur le fonctionnement des enveloppes superficielles, par leur participation à la pédogenèse et à l'altération des roches, notamment. Leurs effets géologiques, à échelle du million d'années, font l'objet de recherches actives, qui s'intéressent tout particulièrement à cette période charnière du Siluro-Dévonien qui voit verdir les continents et, littéralement, « changer la face du monde ».
Les conséquences de l'implantation des végétaux
Les plantes pionnières, cause de la glaciation ordovicienne ?
Aujourd'hui, comme à l'Ordovicien, la végétation pionnière qui colonise un milieu précédemment nu est constituée d'algues et de lichens encroûtants, puis, souvent, de bryophytes (mais parfois fougères ou cactus... selon le climat). Ces organismes s'installent sur des roches et des sédiments en partie altérés par l'activité des microorganismes et participent eux-mêmes à la formation d'un sol, sur lequel ils peuvent s'enraciner et se développer et qu'ils alimentent de leur propres débris organiques.
Néanmoins, jusqu'à récemment, la petitesse de ces mousses et de leurs systèmes de fixation laissait supposer que leur participation à l'altération et à l'érosion des roches était restée modeste dans les premiers temps de la colonisation des terres émergées, au cours de l'Ordovicien. Cette vision a cependant été remise en question récemment par Timothy M. Lenton et ses collègues dans la revue Nature Geoscience (Ref8).
Ces auteurs ont cherché à quantifier expérimentalement l'effet des végétaux non-vasculaires sur l'altération des roches. Leurs expériences ont consisté à placer des grains de roches continentales courantes (granite et andésite) au contact de solutions contenant ou non des mousses (l'espèce actuelle Physcomitrella patens, largement étudié en laboratoire), puis à cultiver ces microcosmes pendant plusieurs heures avant d'y mesurer les quantités de matière organique produites et les teneurs en éléments minéraux dissous (aluminium, calcium, potassium, fer ou encore phosphore). Leurs analyses démontrent que la présence des mousses augmente significativement l'altération chimique des roches (dans le cas de l'andésite, les mousses accroissent les quantités de calcium et de magnésium de près de quatre fois (3,6) et six fois, respectivement ; l'effet sur le granite est moins prononcé, mais net — 1,4 et 1,5 fois, respectivement).
La deuxième étape de leur étude a consisté à introduire dans un modèle numérique préexistant, légèrement modifié, les taux d'altération chimique déduits de leurs expériences pour évaluer leurs effets sur l'évolution d'un système climatique analogue à celui de l'Ordovicien. Le modèle géochimique utilisé, développé par la même équipe (modèle COPSE, Ref2), calcule l'évolution, aux échelles de temps géologiques, des teneurs en carbone, oxygène, phosphore et soufre de l'atmosphère et des écosystèmes émergés et océaniques, en prenant en compte les boucles de rétroaction et les couplages entre les cycles de ces éléments. L'état initial du modèle, mimant la situation de l'Ordovicien inférieur, spécifiait entre autres la teneur atmosphérique en CO2 (estimée entre 14 et 22 fois la teneur actuelle), la température moyenne (plus de 21°C) et la valeur moyenne du δ13C des carbonates marins.
Les données géochimiques disponibles pour cette période ordovicienne montrent :
- un refroidissement des températures globales (déduit des teneurs isotopiques en oxygène de l'apatite de petits fossiles marins, les conodontes) ;
- une baisse du rapport isotopique 87Sr/86Sr de l'eau de mer, signe d'une probable augmentation de l'altération des roches volcaniques, pauvres en 87Sr radiogénique ;
- une chute temporaire du taux de CO2 ainsi que deux pics positifs du δ13C des carbonates à l'Ordovicien supérieur ;
- le développement des dépôts marins de phosphates à la fin de l'Ordovicien ;
- enfin, la glaciation de l'Ordovicien supérieur (glaciation hirnantienne), entre 455 et 440 Ma (figure 8).
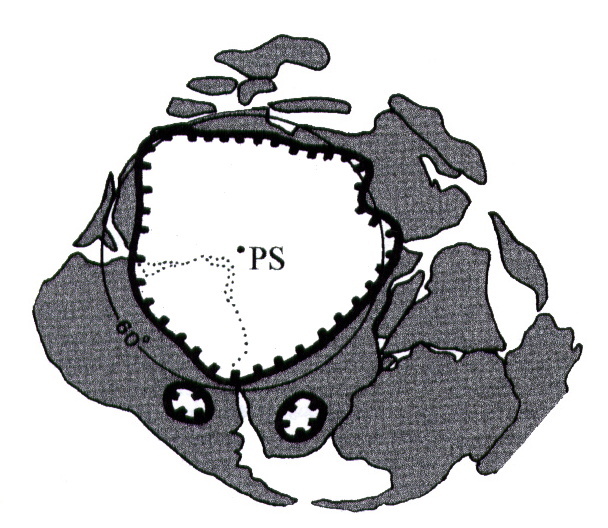
Source - © 1998 in Lethiers (Ref9), d'après Paris et al., 1995
Figure 8. Extension probable de la calotte glaciaire sur le Gondwana à l'Ordovicien terminal
PS : pôle Sud.
Le modèle de Lenton et al. (Ref8) parvient à reproduire ces évolutions du CO2, du δ13C et des températures en simulant la colonisation par les plantes non-vasculaires de 15% des terres actuellement occupées par la végétation en une quinzaine de millions d'années (de 475 à 460 Ma). En particulier, les auteurs obtiennent ainsi une chute du CO2 atmosphérique de 16 à 8 fois la teneur actuelle, du fait du stockage de carbone dans la biomasse et les sols continentaux. Or les études climatiques de l'Ordovicien suggèrent que l'initiation d'une glaciation nécessitait alors un taux de CO2 inférieur ou égal à cette dernière valeur.
De même, ils expliquent les pics de δ13C enregistrés en les attribuant à la libération accrue, mais transitoire, de phosphore continental par les plantes colonisatrices : cet apport de phosphore à l'océan y aurait stimulé la production planctonique, accroissant ainsi l'accumulation de matière organique (pauvre en 13C) dans les sédiments et, en conséquence, le δ13C de l'eau de mer et des carbonates. Ce processus contribuerait également au dépôt de phosphates marins.
Ainsi, ces auteurs concluent que la colonisation des terres émergées aurait, dès ses premières étapes, transformé le fonctionnement des enveloppes terrestres et serait un acteur important des changements climatiques et écologiques qui marquent la fin de l'Ordovicien.
Le développement des forêts et l'extinction du Frasnien-Famennien
Que l'invasion des continents par les végétaux terrestres ait contribué aux bouleversements climatiques et écologiques de l'ère Primaire n'est pas en soi une idée nouvelle, mais elle était jusque là surtout explorée pour les temps dévoniens, marqués par l'expansion des systèmes forestiers.
Ainsi, plusieurs équipes ont cherché à estimer et à modéliser les effets bio-géochimiques du développement des grands végétaux, partant de l'idée que cette augmentation de la taille des plantes s'est accompagnée d'une extension des systèmes racinaires et donc d'un épaississement des sols (figure 9) et d'un enfouissement accru de la matière organique dans cette enveloppe nouvelle.
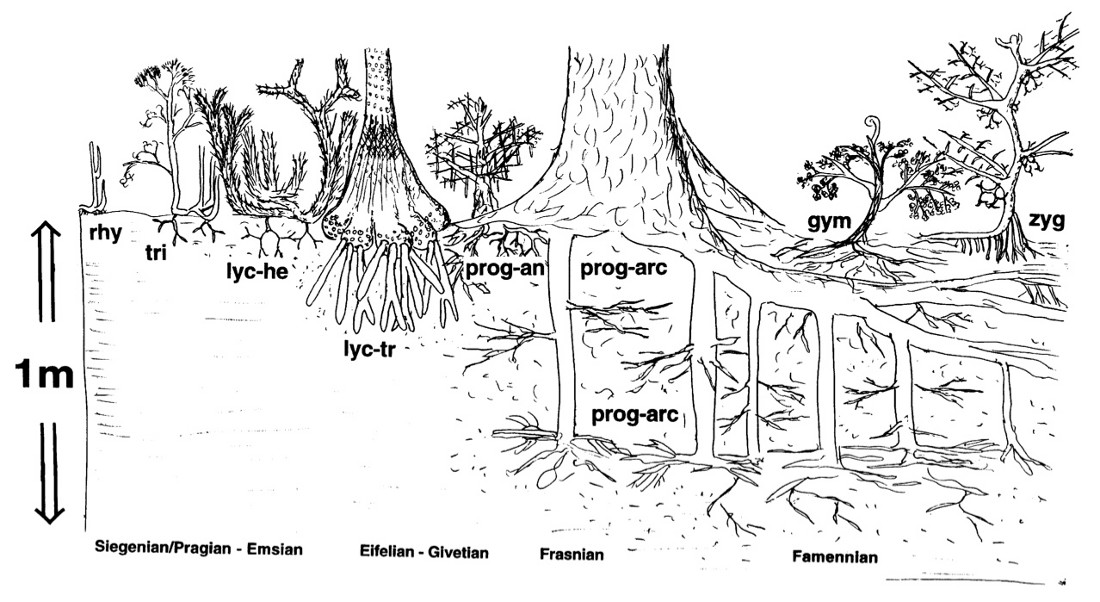
Source - © 1998 Algeo et Schekler (Ref1)
rhy : rhyniophytes (Aglaophyton, Horneophyton) ; tri : trimerophytes (ex : Psilophyton) ; lyc-he : lycopodes (Asteroxylon ou Drepanophycus) ; lyc-tr : lycopodes arborescentes archaïques comme Lepidosigillaria ; prog-an : progymnospermes Aneurophytales, comme Tetraxylopteris ; prog-arc : progymnospermes type Archaeopteris ; gym : gymnospermes archaïques (p. ex. Elkinsia) ; zyg : fougères Zygoptéridales comme Rhacophyton.
Outre cet effet sur le flux de carbone réduit séquestré à long terme dans les sols et les sédiments, les végétaux arborescents modifient les parts relatives de l'altération mécanique et de l'altération chimique des roches : la couverture végétale et les racines altèrent chimiquement les minéraux, mais les protègent de l'entraînement mécanique par les agents d'érosion et stabilisent les sédiments sur lesquels ils se développent. Comme le résume la figure 10, un apport accru de substances chimiques dissoutes à l'océan pourrait, comme à l'Ordovicien, avoir favorisé la prolifération algaire et bactérienne de la zone trophique et l'eutrophisation des plate-formes océaniques, responsable de l'extinction marine qui marque la transition Frasnien - Famennien (derniers étages du Dévonien), il y a 374 Ma. Cette crise, marquée dans les sédiments par plusieurs événements d'anoxie marine, touche principalement le benthos et particulièrement les cnidaires coralliens.
Développement des forêts, climat et paysages du Dévonien
On a vu que l'impact des végétaux arborescents sur les cycles biogéochimiques vient de leur participation à l'altération et à l'érosion des roches, ainsi qu'à leur apport de matière organique aux sédiments. Les conséquences de l'expansion des forêts sur les continents du Dévonien a fait récemment l'objet d'une modélisation numérique par une équipe franco-américaine (Ref7). Ces chercheurs ont couplé un modèle géochimique et climatique à un modèle écologique qui estime la répartition potentielle d'une communauté biologique à partir des cas de présence réellement enregistrés et des caractéristiques écologiques de cet assemblage d'espèces.
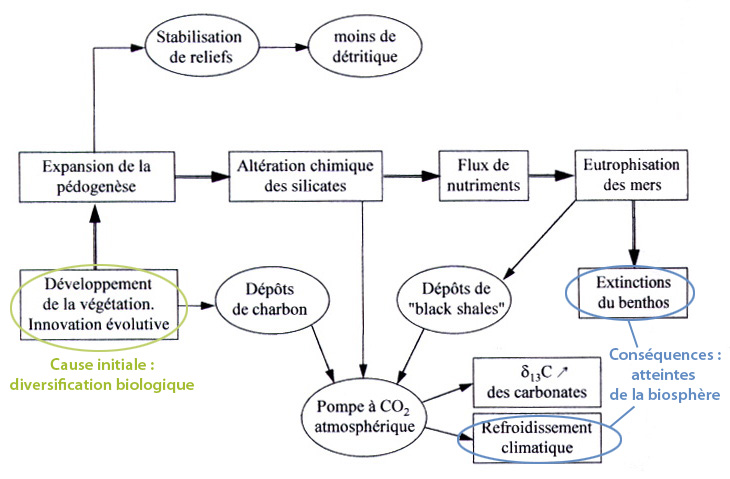
Source - © 1998 Lethiers (Ref9)
Ils ont ainsi utilisé trois grands types de végétation reconnus au cours du Dévonien (communauté archaïque sans formes arborescentes, forêt de Cladoxylopsides arborescentes comme celle de Gilboa, et forêts à arbres ligneux comme Archaeopteris). Les variables écologiques qui contrôlent la distribution de ces communautés végétales sont la température moyenne annuelle de surface et la disponibilité en eau, les trois végétations-types différant par l'extension de leur système racinaire.
L'objectif de cette étude était donc d'évaluer les interactions entre l'effet de l'expansion des végétaux terrestres sur le système climatique et les conséquences rétroactives de ces changements climatiques sur la distribution des végétaux. Le modèle a calculé l'état d'équilibre du système au Dévonien inférieur, moyen et supérieur. Ces calculs indiquent que le développement des végétaux continentaux a accru les taux d'altération des roches silicatées, en particulier dans la zone intertropicale, mais a aussi influencé le ruissellement (les forêts produisent de l'humidité par évapo-transpiration, freinent les masses d'air et favorisent les précipitations). Il a enfin contribué à une nouvelle baisse des pressions partielles de CO2. Cependant le modèle n'obtient pas d'évolution parallèle des taux de CO2 et des températures (la température moyenne globale reste quasi-constante), ce que les auteurs attribuent à une diminution de l'albédo terrestre causé par le verdissement des continents. Dans le même temps, le modèle propose des cartes de la répartition potentielle des communautés végétales considérées : les continents du Dévonien inférieur, il y a 400 Ma, apparaissent encore majoritairement nus, mais sont presque couverts de forêts une trentaine de millions d'années plus tard (figure 11).
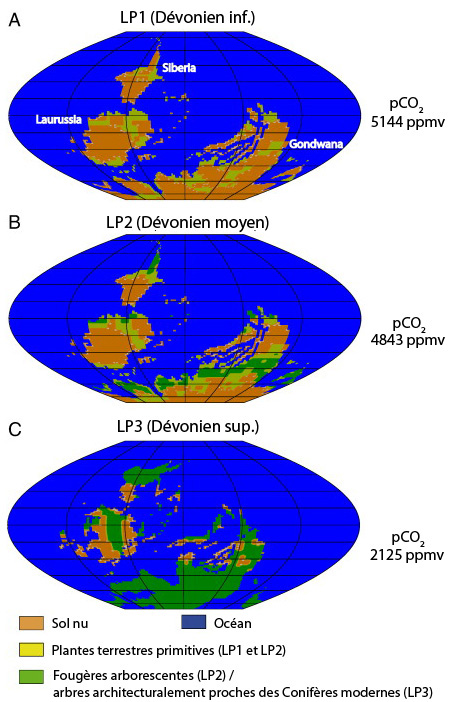
Source - © 2011 Le Hir et al. (Ref7), francisé
Les cartes montrent la répartition potentielle maximale de trois grandes périodes du Dévonien. La végétation, le climat et le cycle du carbone sont à l'équilibre.
Pour finir, cette invasion des terres émergées n'a pas modifié que des chiffres de flux d'éléments chimiques et des bilans géochimiques. Les sédimentologues Martin R. Gibling et Neil S. Davies suggèrent qu'elle a aussi transformé les paysages des régions alluviales ordoviciennes à dévoniennes, Ref5, Ref3.
La morphologie des cours d'eau actuels est contrôlée par la pente, par la stabilité des rives et par les obstacles qui peuvent bloquer les lits : les pentes fortes montrent des rivières en tresses, dont les chenaux multiples changent fréquemment, divaguent et s'anastomosent ; les pentes faibles favorisent la formation de grands méandres.
Mais selon l'analyse de Gibling et Davies, les changements de pente n'expliquent pas la diversification des morphologies fluviales qu'ils reconstituent dans les terrains du début du Phanérozoïque. Ils observent que les rivières et les fleuves du Cambrien sont majoritairement des systèmes en tresses et que l'Ordovicien et le Silurien voient augmenter la fréquence des rivières à méandres et des fleuves au lit stable, parsemé d'îles de grande taille, pérennes, stabilisées par le développement de la végétation. Ce n'est enfin qu'au Dévonien puis au Carbonifère que des amas de troncs emportés par les crues ont pu contribuer à modifier le cours des fleuves en formant des barrages, comme on peut encore l'observer aujourd'hui dans les régions boisées (figure 12).
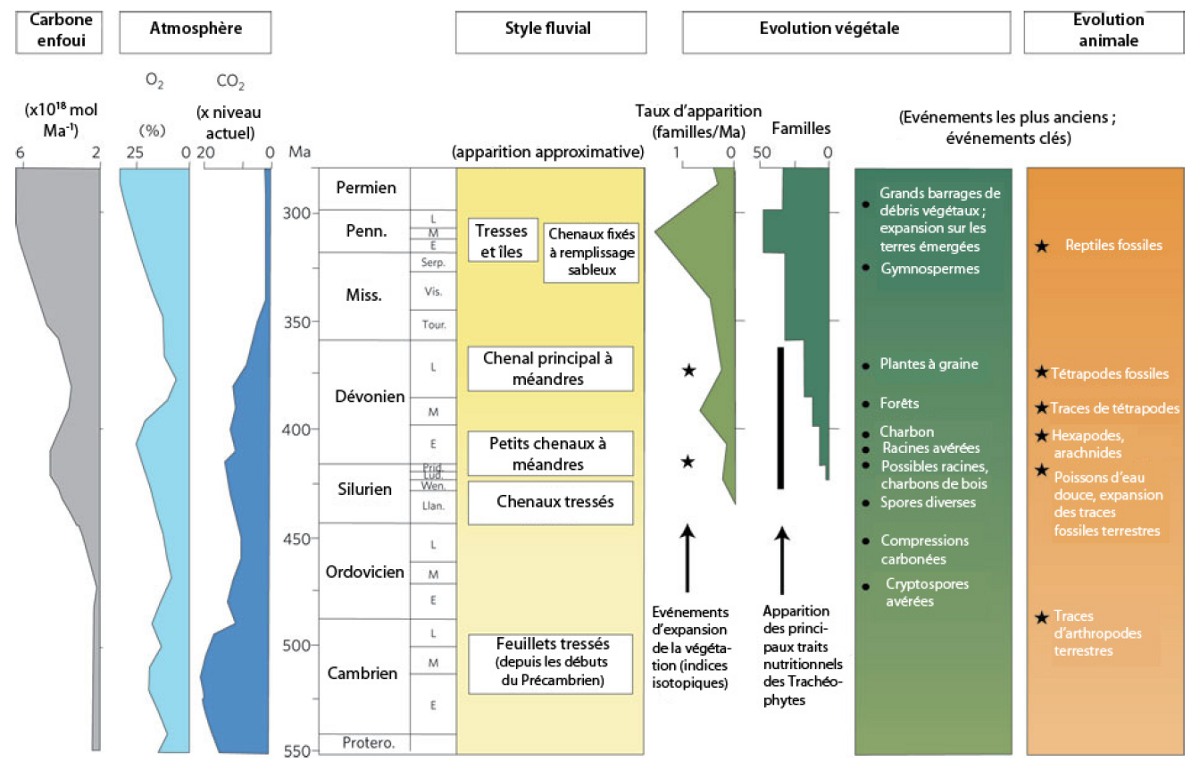
Source - © 2012 Gibling et Davies (Ref5), francisé
Ils identifient également un type de paysage spécifiquement carbonifère, qu'ils qualifient de « plaine alluviale à chenaux fixés », où le lit du cours d'eau est segmenté en larges chenaux, conservant une position stable tandis qu'ils se remplissent de sédiments pour finalement changer abruptement de position (avulsion) (Ref11).
Pour Gibling et Davies (Ref5), cette morphologie spécifique résulte du développement des arbres aux systèmes racinaires bien développés, capables de maintenir les rives des chenaux. Ces auteurs corrèlent donc les changements dans les paysages alluviaux avec l'évolution des végétaux aériens (figure 12). Ils s'appuient d'une part sur les analyses de nombreux affleurements et d'autre part sur les résultats récents des modélisations analogiques, qui réussissent aujourd'hui à étudier les effets du développement végétal sur la stabilité des rives en semant sur leurs modèles des plantes à développement très rapide, comme la luzerne (figure 13).

Source - © 2007-2010 Tal et Paola (Ref17)
Les images A et B sont séparées de 92 jours et 23 cycles de crues.
Conclusion
La « sortie des eaux » des végétaux, entamée à l'Ordovicien, puis la diversification des formes végétales aériennes — dont on n'a présenté ici que quelques étapes — a donc eu de nombreuses répercussions sur les enveloppes externes de la planète. Elle en a modifié les processus sédimentaires, les échanges de matières entre terres émergées et océans, la composition de son atmosphère et par conséquent ses climats, ce qui, en retour, a dû affecter les biocénoses marines et continentales. Le fonctionnement actuel de la biosphère et ses interactions multiples avec les enveloppes externes fluides et solides ne sont par conséquent en place que depuis la fin du Dévonien ou le Carbonifère. Les modifications qu'il a pu connaître depuis paraissent de moindre ampleur — si l'on écarte les épisodes de crises majeures, comme la limite Permo-Trias.
Cet épisode de l'histoire de la Terre est donc unique et irréversible. Son étude est par conséquent d'autant plus utile pour éclaircir les interactions complexes entre biosphère et géosphère, où interviennent de nombreuses boucles de rétroaction.
L'expansion des végétaux a aussi nécessairement eu des répercussions sur l'évolution des organismes qui s'en nourrissent ou s'y abritent. Enfin, même si cette transformation de la surface terrestre a pris des dizaines de millions d'années, elle nous rappelle néanmoins que les modifications anthropiques du couvert végétal peuvent avoir de nombreuses répercussions, difficilement prévisibles, sur le long terme, d'autant plus qu'elles s'effectuent avec une ampleur et à une vitesse jamais vues auparavant.
Quand on interroge des étudiants sur la « sortie des eaux », ils exposent généralement la sortie de Tiktaalik, Acanthostega et Ichthyostega, les premiers tétrapodes connus et leurs ancêtres immédiats. Peu parlent des arthropodes terrestres qui existaient depuis le début de l'Ordovicien, ni de la sortie des eaux des végétaux, objet de cet article. Encore moins parlent de la vraie sortie des eaux, la première, qui a eu lieu dès l'Archéen. En effet, on a des arguments géochimiques indirects et des traces fossiles directes prouvant que des bactéries avaient colonisé les terres émergées dès le début de l'Archéen, sous forme de voiles bactériens (bacterial mats) et autres bio-films.
Références
Algeo, T. J., S. E. Scheckler. 1998. Terrestrial-marine teleconnections in the Devonian : links between the evolution of land plants, weathering processes, and marine anoxic events. Philosophical Transactions of the Royal Society : Biological Sciences, 353(1365) :1471–2970
Bergman, N. M., T. M. Lenton, A. J. Watson, 2004. COPSE : A new model of biogeochemical cycling over Phanerozoic time. American Journal of Science, 304(5) :397–437
Davies, N. S., M. R. Gibling et M. C. Rygel, 2011. Alluvial facies evolution during the Palaeozoic greening of the continents: case studies, conceptual models and modern analogues. Sedimentology, 58(1) :220–258, ISSN 1365-3091
Gensel, P. G., 2008. The Earliest Land Plants. Annual Review of Ecology, Evolution, and Systematics, 39(1) :459–477
Gibling, M. R., N. S. Davies, 2012. Palaeozoic landscapes shaped by plant evolution. Nature Geoscience, 5(2) :99–105
Kenrick, P., C. H. Wellman, H. Schneider, G. D. Edgecombe, 2012. A timeline for terrestrialization: consequences for the carbon cycle in the Palaeozoic. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1588) :519–536
Le Hir, G., Y. Donnadieu, Y. Goddéris, B. Meyer-Berthaud, G. Ramstein, R. C. Blakey, 2011. The climate change caused by the land plant invasion in the Devonian. Earth and Planetary Science Letters, 310(3–4) :203–212, ISSN 0012-821X
Lenton, T. M., M. Crouch, M. Johnson, N. Pires, L. Dolan, 2012. First plants cooled the Ordovician. Nature Geoscience, 5(2)
Lethiers, F.. 1998. Évolution de la biosphère et événements géologiques. Gordon and Breach Science Publishers, Amsterdam
Meyer-Berthaud, B., A. L. Decombeix, 2012. Palaeobotany: In the shade of the oldest forest. Nature, 483(7387) :41–42
Paola, C., 201. Geomorphology: Co-evolution of rivers and plants. Nature Geoscience, 4(9) :583–584
Parry, S., S. Noble, Q. Crowley, C. Wellman, 2011. A high-precision U–Pb age constraint on the Rhynie Chert Konservat-Lagerstätte : time scale and other implications. Journal of the Geological Society, 168(4) :863–872
Selosse, M. A., F. L. Tacon. 1998. The land flora: a phototroph-fungus partnership?Trends in Ecology & Evolution, 13(1) :15 – 20, ISSN 0169-5347
Stein, W. E., C. M. Berry, L. V. Hernick, F. Mannolini. 2012. Surprisingly complex community discovered in the mid-Devonian fossil forest at Gilboa. Nature, 483(7387) :78–81
Stein,W. E., F. Mannolini, L. V. Hernick, E. Landing, C. M. Berry. 2007. Giant cladoxylopsid trees resolve the enigma of the Earth's earliest forest stumps at Gilboa. Nature, 446(7138) :904–907
Tal, M., C. Paola, 2010. Effects of vegetation on channel morphodynamics: results and insights from laboratory experiments. Earth Surface Processes and Landforms, 35(9) :1014–1028, ISSN 1096-9837
Tal, M. et C. Paola, 2007. Dynamic single-thread channels maintained by the interaction of flow and vegetation. Geology, 35(4) :347–350
University of Aberdeen. Rhynie Chert Learning Resource
[1] Le terme anglais chert, qui donne son appellation au site (Rhynie Chert), désigne toutes les formes de roches siliceuses, constituées de silice micro-cristalline (calcédoine ou opale) ou mal cristallisée, d'origine chimique ou biochimique, que le français distinguent par diverses appellations : silex, chailles, accidents siliceux.
[2] Lycophytes ou Lycopodiophytes (Lycopodiophyta) ou Lycopsides (Lycopsida), clade de végétaux vasculaires (Trachéophytes) contenant les actuelles lycopodes, isoètes et sélaginelles.




