Article | 08/11/2017
L'histoire évolutive des iguanes des Galapagos : entre géologie, phylogénie et programmes de conservation
08/11/2017
Résumé
L'évolution des iguanes des Galapagos décryptée grâce aux données de la phylogénie moléculaire avec une interprétation de cette histoire évolutive en fonction de la géologie des iles. Les programmes de conservation des populations d'iguanes aux Galapagos.
Table des matières
Un article sur ce sujet est paru dans le magazine Espèces, n°24 de juin-août 2017. Il a été co-écrit avec Thibault Lorin, dont les judicieuses remarques ont permis de grandement améliorer la base ayant servi à la version magazine pour aboutir au présent article, repensé et étoffé pour Planet-Terre.
Introduction
Les iles Galapagos (galápago = tortue terrestre en espagnol d'Amérique latine) sont considérées comme un véritable laboratoire naturel de l'évolution. De nombreuses espèces endémiques bien connues du grand public ont depuis longtemps attiré l'attention des scientifiques.
 Source - © 2004 Pierre Thomas | |
 Source - © 2005 Jean Blanchard |  Source - © 2004 Pierre Thomas Figure 3. Un accouplement de tortues géantes des Galapagos Les individus ne sont matures sexuellement qu'à l'âge de 20 à 25 ans. La vitesse de reproduction de ces animaux est donc très lente, ce qui fragilise ces populations. Toutes les espèces de tortues géantes sont d'ailleurs sur la liste rouge de l'UICN. |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 4. Un pinson de Darwin aux Galapagos |  Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 5. Un iguane marin aux Galapagos |
C'est ainsi qu'au nom des Galapagos résonne celui de Charles Darwin, le co-découvreur de la sélection naturelle, l'un des mécanismes de l'évolution. C'est après avoir observé les pinsons des Galapagos, lors de son voyage sur le HMS Beagle en 1835, que Darwin élabora sa théorie (cf. Sélection naturelle, VIH, pinsons : ce que Darwin ne pouvait pas savoir). Pourtant, si on revient à ses écrits et plus particulièrement aux notes qu'il a prises pendant son séjour aux Galapagos, on se rend compte que les pinsons n'étaient alors pas du tout son sujet d'observation favori : il leur consacre à peine une page ! Sa curiosité est alors tournée vers d'autres insulaires, bien plus « hideux » et « stupides » de ses propres mots, les iguanes des Galapagos.
Darwin leur consacrera plus de six pages dans les notes qu'il publiera à son retour [L1], il ne peut alors s'empêcher de montrer toute sa fascination et sa curiosité pour ces animaux qui font, selon lui, parti d'un « genre remarquable de lézards (sic) […] particulier à cet archipel ». Ainsi il écrivait : « Il est fort intéressant, en somme, de trouver un genre bien caractérisé possédant une espèce marine et une espèce terrestre, et confiné dans une si petite partie du monde ».
Darwin parlait bien sûr des iguanes des Galapagos qu'on classe aujourd'hui dans deux genres bien distincts : les iguanes terrestres (Conolophus, trois espèces) et les iguanes marins (Amblyrhynchus, une espèce et 11 sous-espèces). Ce dernier genre était d'ailleurs pour lui « de loin l'espèce (sic) la plus remarquable ». Malgré toutes les observations qu'il consignera sur ces iguanes, il ne semble pas qu'il ait cherché à savoir comment ces deux genres se sont retrouvés dans cette « si petite partie du monde ». Une question qui a le mérite de se poser dans le contexte de la théorie de l'évolution qu'il développera bien des années après.
Ce sont donc des travaux récents, poursuivant des recherches initiées dans les années 1980, qui nous éclairent sur ce sujet. Nous proposons de les mettre en lien avec la géologie et la géographie des Galapagos, donc ses contraintes environnementales, pour mieux comprendre l'origine évolutive de ces passionnants iguanes. Et c'est en comprenant leur histoire que nous pourrons appliquer des programmes de conservation qui protégeront efficacement leur diversité.
Géographie et géologie des iles Galapagos
Les Galapagos, un archipel d'iles équatoriales dans le Pacifique
Les Galapagos forment un ensemble d'une centaine d'iles et d'ilots du Pacifique. Elles s'étalent du Nord-Ouest au Sud-Est sur près de 400 km. L'ile la plus orientale, San Cristobal, est à 960 km des côtes de l'Équateur, pays auquel cet archipel est rattaché sous le nom de « province des Galapagos ».
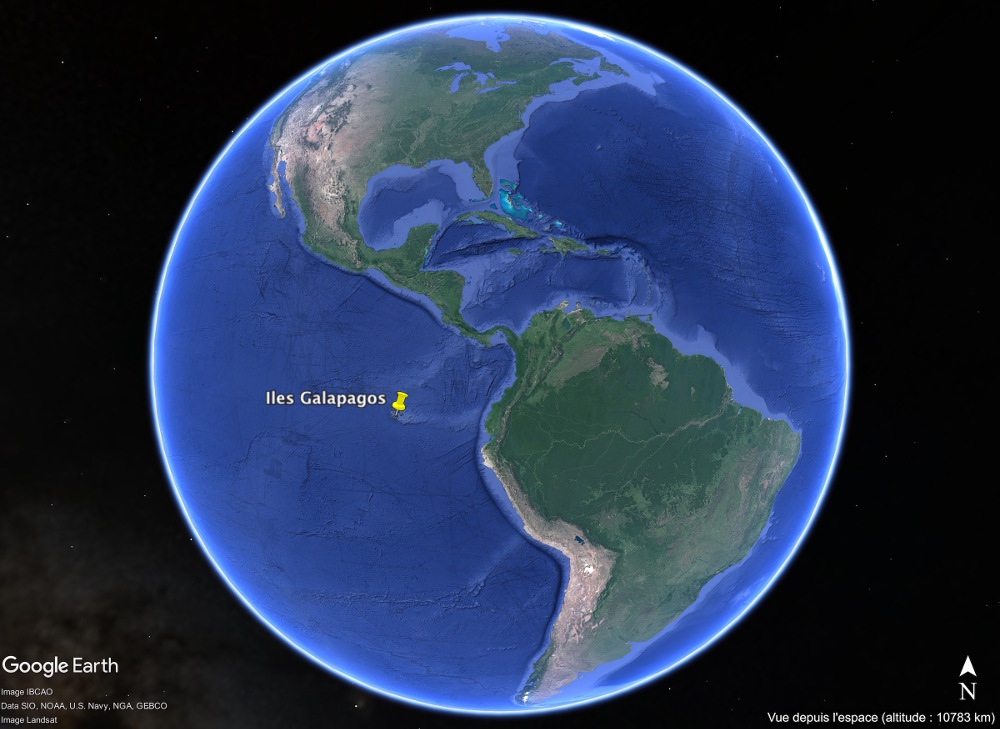 Source - © 2017 Google Earth | 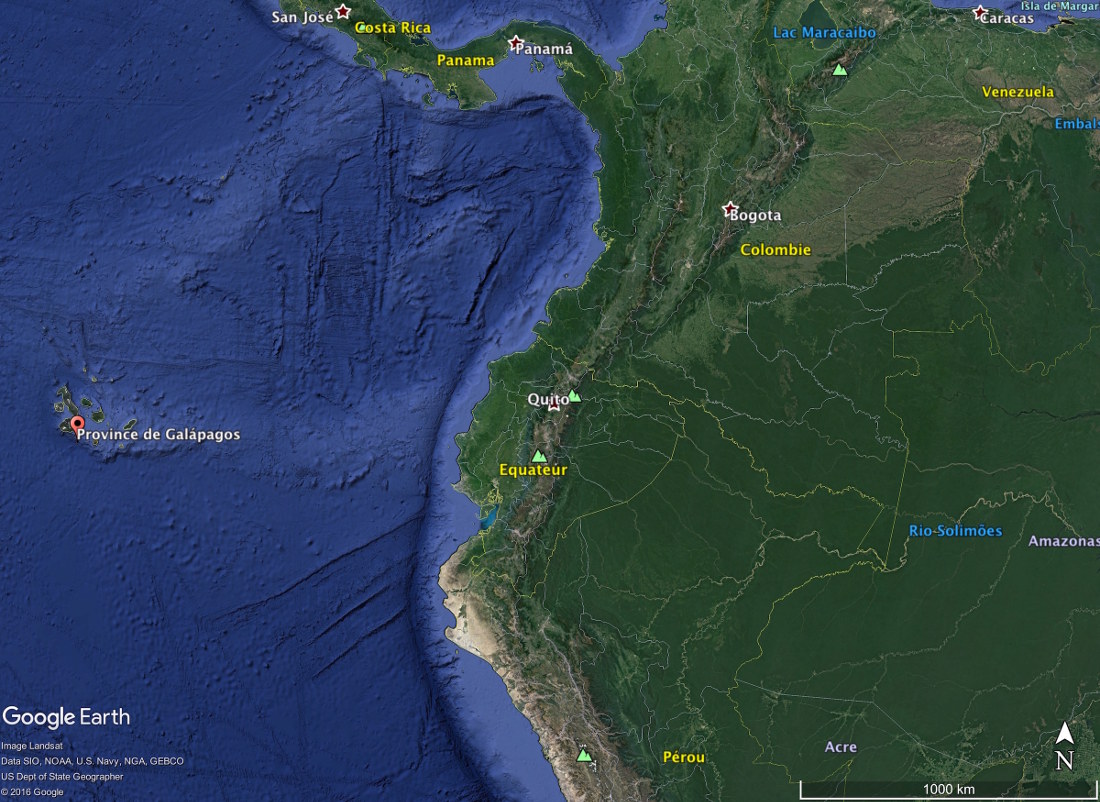 Source - © 2017 Google Earth Figure 7. Localisation des Galapagos au large des côtes équatoriennes Les iles Galapagos se situent à près de 1 000 km de la côte équatorienne. La ride de Carnegie, "trace" du point chaud des Galapagos, se suit à l'Est de l'archipel jusqu'à la fosse de subduction. |
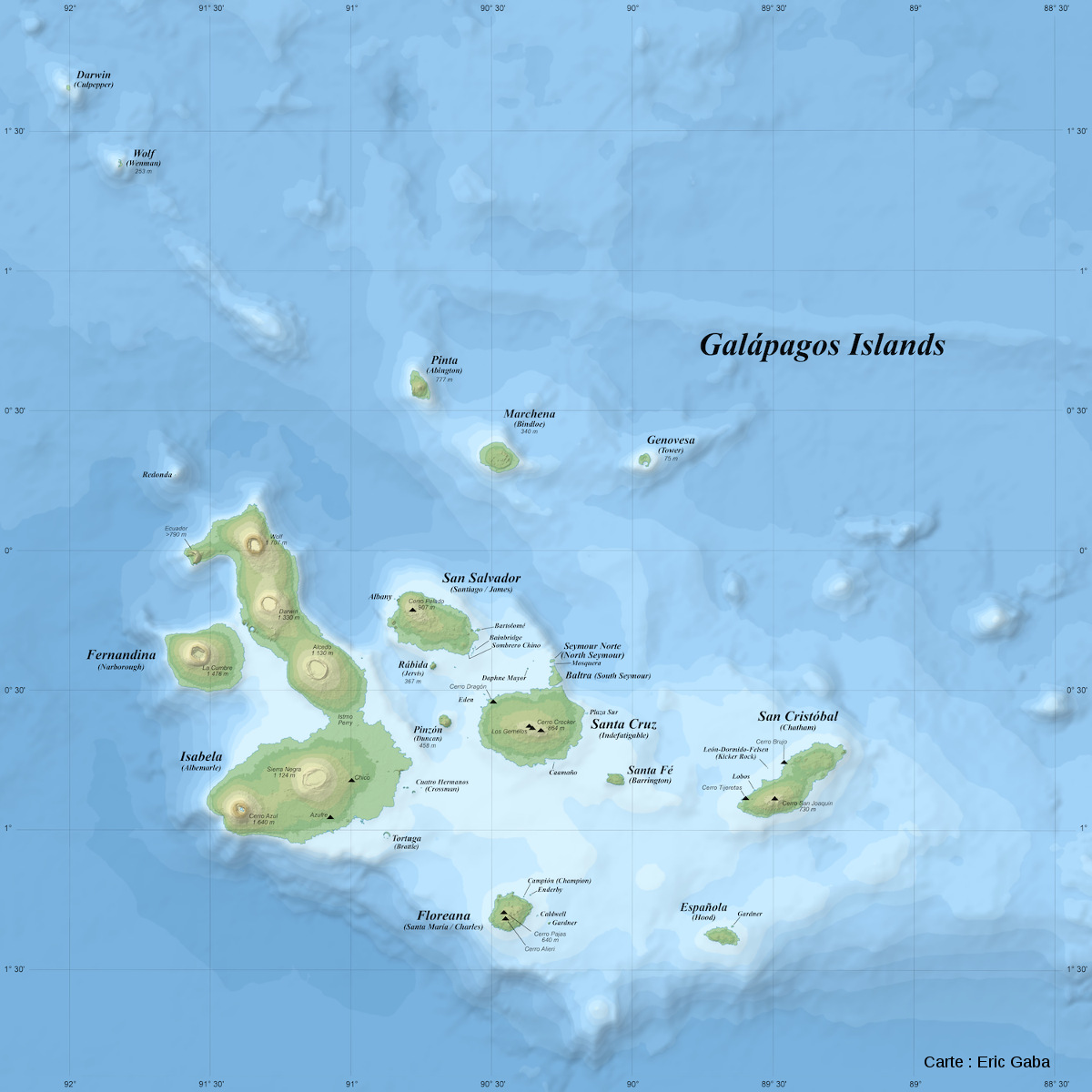 Source - © 2017 Eric Gaba, CC BY-SA 4.0 | |
Les Galapagos, la manifestation en surface d'un point chaud
La vingtaine d'iles majeures des Galapagos est formée de plusieurs centaines de volcans, dont certains sont actifs aujourd'hui et entrent régulièrement en éruption. La topographie de chaque ile nous renseigne sur son activité volcanique et donc, indirectement, sur son âge. Les iles Isabela et Fernandina, à l'Ouest, possèdent des reliefs qui culminent à 1707 et 1476 m respectivement, alors que San Cristobal, à l'Est, culmine à 730 m. Cette dernière ile n'est plus active depuis plusieurs siècles (Geist et al., 1986 [6]). Les laves émises aux Galapagos et les roches correspondantes sont majoritairement basaltiques. Ces volcans majeurs sont de type volcan bouclier (cf. Exemples de volcans boucliers : Galapagos, La Réunion, Islande, Sicile, Tahiti).
Ces observations s'expliquent bien par un modèle de plaque superficielle mobile "percée" par un point chaud "fixe". Les Galapagos sont, comme la Réunion ou Hawaii, issues du fonctionnement d'un point chaud enraciné profondément dans le manteau terrestre et sans doute jusque vers l'interface manteau/noyau. Celui-ci se situe en dessous d'une des plaques du Pacifique, la plaque de Nazca. Cette plaque se forme depuis une dorsale à l'Ouest et plonge dans le manteau à l'Est au niveau de la cordillère des Andes. Elle se déplace ainsi de plus de 6 cm par an vers l'Est. Le point chaud étant supposé fixe, son volcanisme itératif, mais régulier depuis une dizaine de millions d'années, permet alors de former des iles qui se déplacent vers l'Est au fur et à mesure qu'elles vieillissent. La datation des différentes iles (en particulier par radiochronologie sur les basaltes) conforte ce modèle (tableau de données ci-dessous, Geist et al., 2014 [7]). À l'Est de la dernière ile, cette activité ancienne se manifeste par une ride asismique, la ride de Carnegie, équivalent "galapagosien" de la ride asismique de l'Empereur vis-à-vis du point chaud d'Hawaii. La situation est en fait plus complexe à cause de la naissance récente de la dorsale Sud-Coco qui a sans doute séparé la trace du point chaud en deux rides asismiques (Carnegie et Coco).
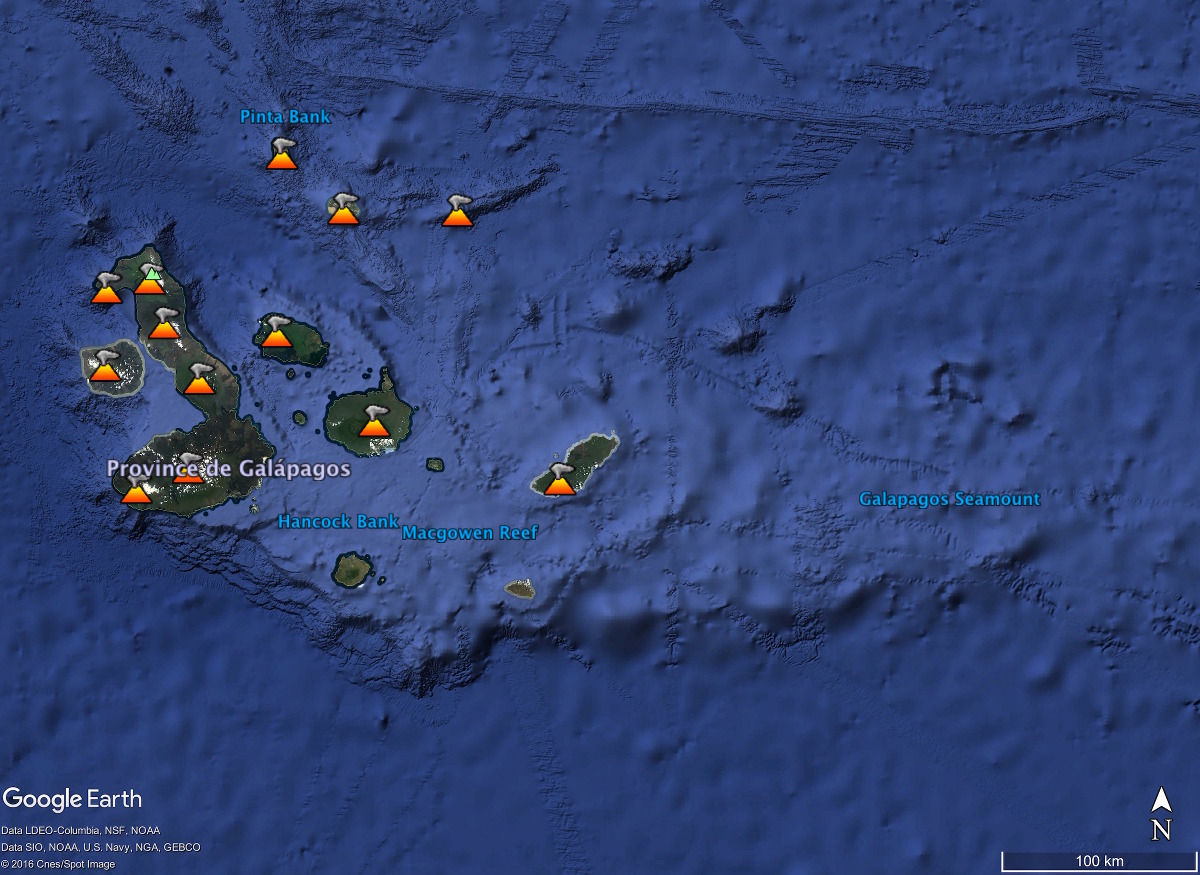 Source - © 2017 Google Earth Figure 9. Les Galapagos, des iles volcaniques issues du fonctionnement d'un point chaud La plupart des volcans des Galapagos sont en activités, les plus orientaux sont endormis. Les iles les plus anciennes se situent à l'Est, les plus récentes à l'Ouest. | |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Ce que les géologues locaux ont d'abord appelé « tuf » correspond en fait à des dépôts phréatomagmatiques de type surtseyien. Les cratères de tuf se sont formés sous l'eau lors du contact brutal entre les cendres chaudes et l'eau froide. Ceci souligne bien l'origine marine de ces iles. |  Source - © 2015 Florent Figon - CC BY-SA 3.0 |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 12. Le cratère de « tuf » surtseyien de l'ile Daphne Major Daphne Major est un laboratoire naturel de l'évolution. Le couple de chercheurs Grant y ont plus particulièrement étudié l'évolution des populations de pinsons de Darwin pendant plus de 30 ans. |  Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 13. Des spatter cones, des laves cordées et un tunnel de lave sur l'ile Bartolome Vous pouvez retrouver des explications sur ces spatter cones dans Les spatter cones et les hornitos, des édifices volcaniques à pentes raides mais pourtant engendrés par des laves très fluides. |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 14. L'immense caldeira et le toit de lave du volcan Sierra Negra au Sud de l'ile Isabela Ce cratère fait plus de 100 km2 et sa dernière éruption remonte à 2005. | |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 15. Cônes adventifs et coulées pahoehoe sur l'ile Santiago Vous pouvez retrouver d'autres exemples de cônes adventifs dans Cônes adventifs sur les flancs du Pico (volcan des Açores), dont l'un a été aménagé en enclos à bétail, et de coulées pahoehoe dans Laves « en tripes » (entrail pa hoe hoe lava flow), Piton de la Fournaise, île de la Réunion. |  Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 16. Los tuneles, tunnels de lave côtiers effondrés et en partie immergés au Sud de l'ile Isabela Voir aussi d'autres tunnels dans Lucarnes ( skylight ) et formation des tunnels de lave ( lava tube ). |
Ile (d'Ouest en Est) | Date d'émergence (en millions d'années) | Point culminant (en mètres) |
|---|---|---|
Fernandina | 0,032 – 0,06 Ma | 1476 m (Volcan La Cumbre) |
Isabela | 0,5 – 0,8 Ma | 1707 m (Volcan Wolf) |
Santa Cruz | 1,1 – 2,3 Ma | 864 m (Volcan Cerro Crocker) |
Española | 3,0 – 3,5 Ma | 206 m |
San Cristobal | 2,4 – 4,0 Ma | 730 m (Volcan Cerro San Joaquin) |
L'une des prédictions de ce modèle est que nous devrions retrouver des iles immergées ou a minima des laves basaltiques issues du fonctionnement du point chaud il y a plus de 4 Ma. Des études récentes montrent effectivement qu'il existe un plancher basaltique à l'Est de San Cristobal ; on peut d'ailleurs le voir sur des cartes bathymétriques et topographiques. C'est ce qu'on appelle les Galapagos Seamount (les montagnes sous-marines des Galapagos) avec des reliefs faisant penser à d'anciennes iles. Elles sont aujourd'hui immergées à cause de l'érosion et de la subsidence thermique de la lithosphère océanique qui s'éloigne de la dorsale. Ces vestiges d'iles ont pu être datés et remontent à près de 11 Ma (Merlen, 2014 [12]). La ride asismique de Carnegie prolonge ce dispositif vers l'Est comme la ride de l'Empereur prolonge l'archipel hawaiien.
Le climat des Galapagos
Alors que les Galapagos sont à cheval sur l'équateur, le climat y est plutôt sec. La station de la Fondation Charles Darwin sur Santa Cruz enregistre des précipitations annuelles moyennes autour de 300 mm. Cette sécheresse inhabituelle sous l'équateur est due au fait que les iles Galapagos sont baignées par un bras Nord du célèbre courant froid de Humbolt (également appelé courant du Pérou). Ces eaux froides ne sont sans doute pas étrangères à l'abondance de pinnipèdes et de manchots sur ces plages équatoriales. À l'inverse, lors des épisodes El Niño, ces précipitations atteignent près de 3000 mm à l'année ! Étant loin de la côte, donc préservées de toute influence continentale, le climat des iles Galapagos est directement dépendant des courants marins, d'où l'importance des phénomènes El Niño.
Le climat et la végétation d'une ile des Galapagos varient en fonction de sa topographie et de son activité volcanique. Une même ile, selon la position de son point culminant (effet de fœhn) et la date de sa dernière éruption, peut arborer une côte à la végétation luxuriante et une côte à l'environnement désertique. Cette dynamique temporelle et spatiale du climat, que nous détaillons dans les paragraphes suivants, a de profondes implications pour l'évolution des êtres vivants qui l'habitent.
Les écosystèmes des iles anciennes
Comme on l'a vu précédemment, lorsqu'une ile est ancienne, son climat et sa végétation sont très dépendantes de sa géographie et de sa géomorphologie.
Prenons l'exemple de San Cristobal. Cette ile âgée de 2,4 à 4 Ma est globalement découpée en deux zones de végétations très différentes : le Sud et le Nord. La côte Sud est baignée par des courants marins et des vents qui amènent de l'humidité et donc des précipitations. Une végétation de type intertropicale s'y développe alors. Pour ce qui est de la partie Nord de l'ile, que les coulées basaltiques soient plus ou moins récentes, les quelques massifs volcaniques culminant à 700 m (Cerro San Joaquin, la lagune El Junco...) suffisent pour arrêter les masses d'airs humides et ainsi empêcher les précipitations d'atteindre le Nord de l'ile. Le climat y est beaucoup plus sec, et la végétation est dominée par des plantes grasses et des herbes.
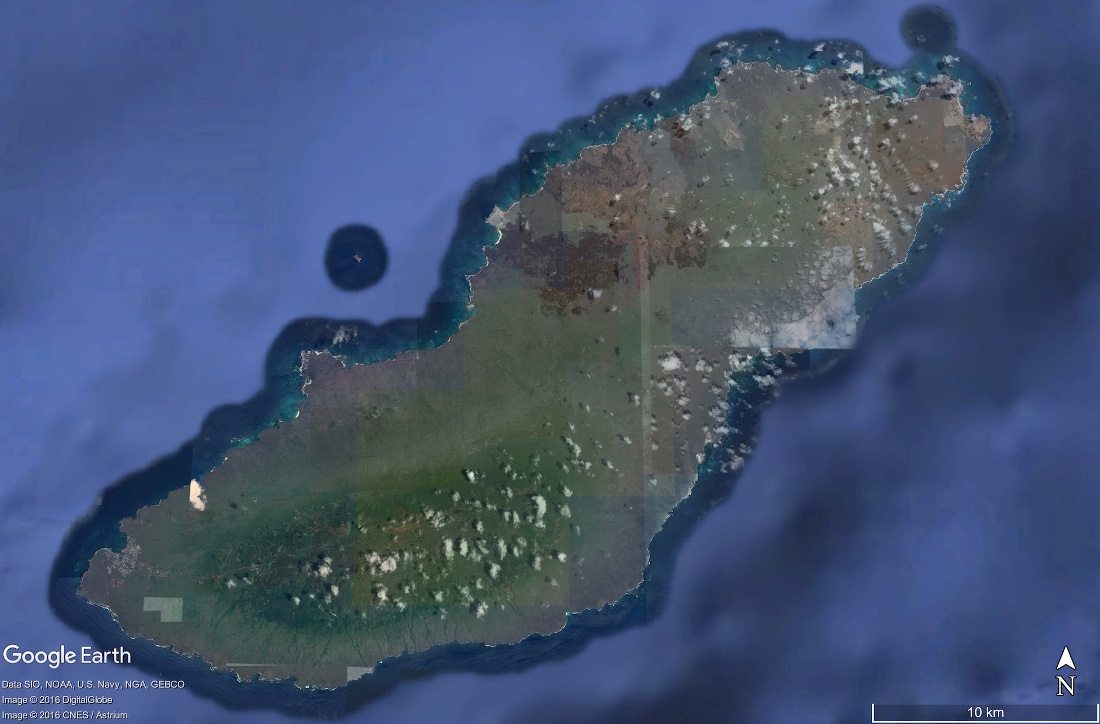
Source - © 2017 Google Earth
Figure 17. Vue satellite de l'ile San Cristobal et de sa végétation, Galapagos
Cette vue satellite montre bien la différence de végétation entre le côté Nord et le côté Sud des iles des Galapagos. Le Sud est beaucoup plus humide, et donc verdoyant, que le Nord. De plus, le relief arrête les nuages et favorise donc les précipitations sur ce même côté de l'ile.
Cet exemple s'inscrit dans le modèle des zones de végétation des iles Galapagos. Nous ne traiterons pas de chaque zone, mais les zones les plus humides (écozones à Scalesia, Astéracée arboresente endémique, et à Miconia, Mélastomatavée arborescente) ne peuvent se mettre en place que sur des reliefs hauts et humides. Les zones restantes sont beaucoup plus sèches.
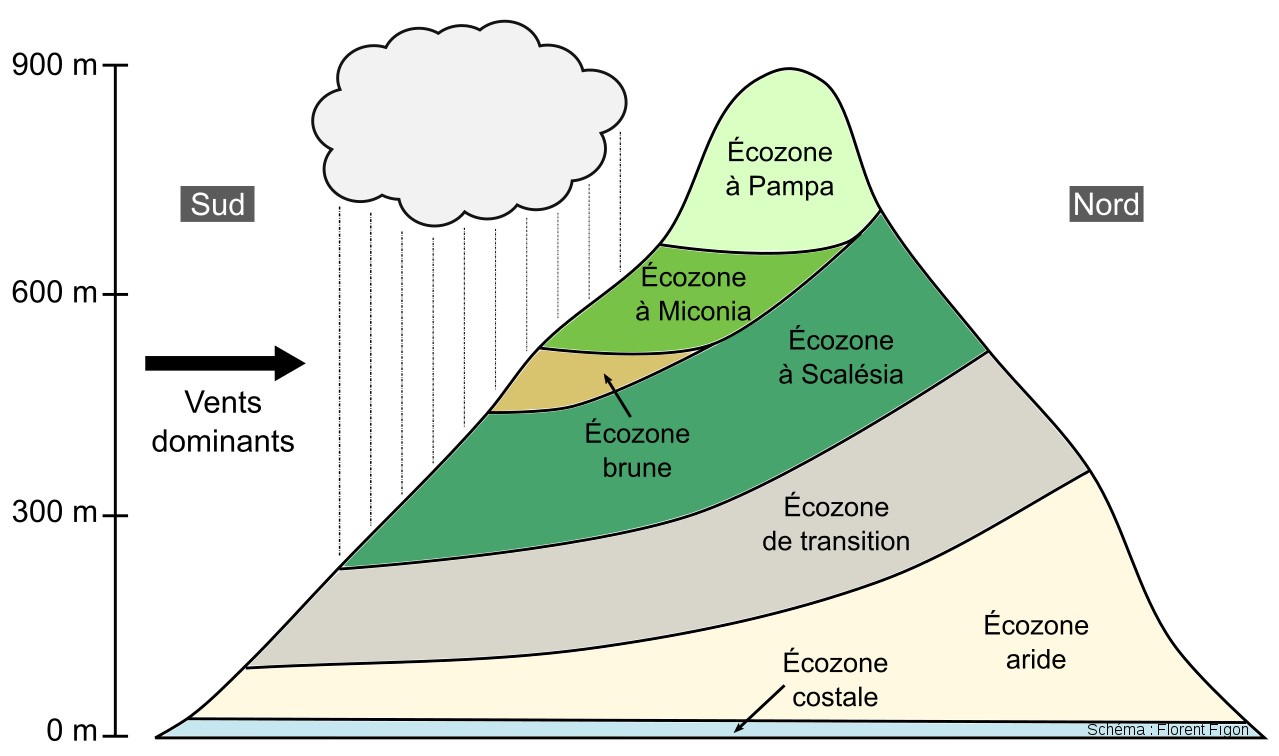
Source - © 2015 Florent Figon - CC BY-SA 3.0
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 19. Passage de la zone costale à la zone aride sur l'ile San Cristobal L'écozone costale est la première bande végétale visible sur cette photo. Elle est principalement constituée de plantes grasses résistantes à la dessiccation et aux hautes teneurs en sels, et parfois de mangroves. La zone aride se situe sur les pentes en arrière-plan. Celle-ci est moins dense en végétaux. | |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 20. Passage des zones arides et/ou de transition à la zone à Scalesia sur l'ile San Cristobal On reconnait en arrière-plan la zone aride peu végétalisée. Une transition se fait alors vers une végétation plus dense et verdoyante au premier plan, la zone à Scalesia. |  Source - © 2005 Jean Blanchard Figure 21. Zone aride sur l'ile Isabella Cette zone où l'on retrouve des cactus du genre Opuntia est typiquement le milieu de vie des iguanes terrestres qui se nourrissent de leurs fruits et de leurs "raquettes". |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 22. Zone à Scalesia sur l'ile Santa Cruz Dans cette zone à la végétation dense, pas d'iguanes, mais de nombreuses espèces d'oiseaux, des tortues terrestres, etc. |  Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 23. Zone à Miconia sur l'ile San Cristobal Cette zone est particulièrement importante pour la nidification des pétrels, des oiseaux marins qui vont se nourrir à la surface des mers à plusieurs kilomètres des côtes. La zone à Miconia est particulièrement menacée par des plantes introduites et invasives, telle que la ronce. |
Les écosystèmes des iles récentes
Pour les iles plus récentes, le contrôle des écosystèmes se fait beaucoup plus par l'activité volcanique que par des effets climatologiques (le volcan Sierra Negra fait exception avec ses pentes Sud très humides et arborées et ses pentes Nord quasi désertiques). Selon la lithologie, seules quelques plantes bien spécifiques arrivent à se développer. Dans le cas de l'ile Bartolome, les cendres et « tufs » sont colonisées par les espèces pionnières Tiquilla (Boraginacée) et Chamaesyce (Euphorbiacée), alors que les laves le sont par le cactus Brachycereus.
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 24. Plantes pionnières du genre Tiquilia colonisant les cendres et tufs de l'ile Bartolome Ces végétaux sont les premiers à coloniser une ile ou une nouvelle région d'une ile suite à une éruption volcanique. Leur faible densité et le fait qu'ils ne soient pas une nourriture de choix pour les iguanes (qui les disperseraient alors) ne permet pas à des populations importantes de s'y installer de manière durable. Voir ou revoir à ce propos La colonisation des coulées de lave par des cactus en climat chaud et sec, iles de Fernandina et de Bartolomé, archipel des Galapagos (Équateur). | |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Ces végétaux sont les premiers à coloniser une ile ou une nouvelle région d'une ile suite à une éruption volcanique. Leur faible densité et le fait qu'ils ne soient pas une nourriture de choix pour les iguanes (qui les disperseraient alors) ne permet pas à des populations importantes de s'y installer de manière durable. Voir ou revoir à ce propos La colonisation des coulées de lave par des cactus en climat chaud et sec, iles de Fernandina et de Bartolomé, archipel des Galapagos (Équateur). |  Source - © 2015 Florent Figon - CC BY-SA 3.0 Figure 26. Brachycereus nesioticus, un cactus pionnier des champs de lave de l'ile Bartolome Ces végétaux sont les premiers à coloniser une ile ou une nouvelle région d'une ile suite à une éruption volcanique. Leur faible densité et le fait qu'ils ne soient pas une nourriture de choix pour les iguanes (qui les disperseraient alors) ne permet pas à des populations importantes de s'y installer de manière durable. Voir ou revoir à ce propos La colonisation des coulées de lave par des cactus en climat chaud et sec, iles de Fernandina et de Bartolomé, archipel des Galapagos (Équateur). |
Connaître le climat, et donc la végétation, d'une ile est important si on veut comprendre comment les iguanes des Galapagos ont évolué en les formes que nous connaissons actuellement. En effet, une ile selon son âge, son altitude, son activité volcanique... imposera des contraintes à la végétation qui y pousse et donc in fine aux iguanes qui la coloniseront.
Les iguanes des Galapagos : des espèces endémiques aux modes de vie variés
Les iguanes terrestres
Tous les iguanes vivent dans la région de l'Amérique du Sud et de l'Amérique centrale. Mis à part les iguanes marins des Galapagos, ils sont tous herbivores et insectivores. Trois espèces endémiques d'iguanes terrestres sont présentes aux Galapagos : Conolophus subcristatus, C. pallidus (de l'ile Santa Fe) et C. marthae (l'iguane rose découvert en 1986 et nommé seulement en 2009 !). Les individus de ces trois espèces se distinguent principalement par la couleur de leurs écailles. L'archipel compte jusqu'à 10 000 individus d'iguanes terrestres avec une très large majorité de C. subcristatus.
 Source - © 2005 Alain Campo-Paysaa |  Source - © 2013 Benjamint 444 - GFDL 1.2 |
 Source - © 2009 Post of Ecuador | |
Ces iguanes peuvent mesurer jusqu'à 1 m et peser près de 13 kg. Les adultes sont majoritairement herbivores ; ils se nourrissent des cactus du genre Opuntia en mangeant aussi bien les parties végétatives (les pads ou raquettes) que les fruits. Les juvéniles sont insectivores (tout comme les lézards des laves, un autre groupe d'espèces endémiques de ces iles) ; ils deviennent phytophages en grandissant. On retrouve ces iguanes dans les terres, où ils nidifient en creusant des terriers, jamais sur les côtes.
 Source - © 2013 Benjamint 444 - GFDL 1.2 | |
 Source - © 2005 Jean Blanchard |  Source - © 2005 Jean Blanchard |
L'iguane marin
Bien que moins connu que les pinsons de Darwin ou les tortues géantes des Galapagos, l'iguane marin (Amblyrhynchus cristatus) est une espèce emblématique de ces iles à plusieurs titres. Elle est non seulement unique parce qu'elle est endémique de ces iles, mais surtout elle est la seule représentante de son groupe à avoir un mode de vie amphibien (ou semi-aquatique) ! Son alimentation, à l'âge adulte, est uniquement composée d'algues rouges et vertes qu'elle trouve sur les fonds marins et dans les zones intertidales. Elle est donc inféodée aux côtes, que ce soit des plages ou des falaises. Les mâles vont plus souvent se nourrir en s'éloignant de quelques mètres des côtes et en plongeant à plusieurs mètres de profondeur. Les femelles et les juvéniles se nourrissent plus souvent sur les rochers battus par l'eau et dans les zones intertidales.
Si une seule espèce semble exister, elle se subdivise en plusieurs sous-espèces selon les iles. On en compte aujourd'hui 11 (Miralles et al., 2017 [11]). Ces sous-espèces varient principalement par leur taille (de 0,75 à 1,3 m) et leur couleur (du noir profond au noir tacheté de rouge et de vert).
 Source - © 2015 Florent Figon - CC BY-SA 3.0 Le basking est un comportement permettant à l'animal de se réchauffer après être allé plonger pour se nourrir. |  Source - © 2015 Florent Figon - CC BY-SA 3.0 La différence de couleur de ce mâle par rapport à celui de la photo précédente est due à la saison de reproduction. Les iguanes mâles arborent alors une livrée jaune-rouge qui pourrait attirer les femelles. |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 |  Source - © 2015 Florent Figon - CC BY-SA 3.0 |
 Source - © 2015 Florent Figon - CC BY-SA 3.0 | |
 Source - © 2005 Jean Blanchard |  Source - © 2005 Jean Blanchard |
 Source - © 2004 Jean-Marc Moullet L'iguane marin et le crabe Sally-pied-léger vivent dans les mêmes milieux. Ce crabe est, lui aussi, emblématique des Galapagos. |  Source - © 2004 Jean-Marc Moullet L'iguane marin et le crabe Sally-pied-léger vivent dans les mêmes milieux. Ce crabe, lui aussi emblématique des Galapagos, mange les peaux mortes et certains parasites (tiques notamment) présents sur les iguanes marins. Ces derniers ne sont nullement incommodés et semblent même profiter de ces séances de toilettage ! Un bel exemple de commensalisme voire de mutualisme à bénéfices réciproques, qui implique une coévolution comportementale. |
Source - © 2015 Florent Figon - CC BY-SA 3.0
Vidéo d'un iguane en train de se nourrir sous l'eau.
L'iguane marin possède diverses adaptations en lien avec son mode de vie original (voir tableau ci-dessous).
Type d'adaptation | Adaptation | Avantage conférée en milieu marin |
|---|---|---|
Morphologique | Pattes légèrement palmées | Augmentation de la vitesse de nage |
Morphologique | Doigts griffus longs et puissants | Meilleure accroche aux roches |
Morphologique | Face aplatie | Permet de brouter les algues fixées aux rochers |
Morphologique | Queue aplatie latéralement | Stabilisation de la nage |
Morphologique | Corps sombre | Diminution de l'albédo permettant un réchauffement plus rapide par absorption des rayons lumineux (les iguanes perdent jusqu'à 10°C pendant la plongée) |
Physiologique | Diminution drastique de la fréquence cardiaque en plongée | Optimisation de l'oxygénation en plongée |
Physiologique | Excrétion de sels de chlorure et de potassium par les narines | Accommodation de la teneur élevée en sel de l'eau de mer et des algues ingérées |
Comportementale | Comportement grégaire | Profiter de la chaleur des congénères |
Comportementale | Position de basking | Optimisation du réchauffement du corps en variant l'inclinaison de la tête par rapport aux rayons du soleil et au substrat rocheux brulant (atteignant jusqu'à 60°C !) ; la température corporelle doit atteindre la température critique de 35°C. |
Comportementale | Commensalisme / Mutualisme avec le crabe Sally-pied-léger | Nettoyage des tiques accrochées aux iguanes et diminution de la pression parasitaire |
L'ensemble des données concernant ces deux genres d'iguanes aux modes vies opposés pose la question de leur histoire évolutive et donc de leur origine. Sont-ils très apparentés ? Proviennent-ils d'une même population d'iguanes qui ont colonisé les Galapagos ? Comment cette population a-t-elle pu atteindre des iles si éloignées ? Quand et combien de fois cela s'est-il produit ?
L'origine des iguanes des Galapagos
Comment coloniser des iles si lointaines ?
Le problème de la possibilité de coloniser de telles iles se pose à partir des observations suivantes :
- les Galapagos sont à près de 1000 km des côtes Sud-américaines ;
- le fait que le point chaud ne fonctionne que depuis quelques dizaines de millions d'années implique que ces iles n'ont pas pu être en contact avec les continents dans le passé ;
- la profondeur de l'océan entre les iles et la côte ne permet pas d'envisager un « pont émergé » pendant de grandes phases de régression marines (la connexion entre les iles est par contre très probable) ;
- la faune et la flore terrestres de ces iles sont très différentes, en termes de groupes de taxons présents et même abondants, de la faune et de la flore terrestre continentale (tableau ci-dessous) ; on ne prendra pas en compte les espèces introduites par l'homme qui ont pu bénéficier de leurs moyens de transports depuis la découverte des iles en 1535.
Groupes de taxons terrestres | Abondance aux Galapagos par rapport à l'abondance mondiale (ne prend pas en compte les espèces introduites par l'homme) |
|---|---|
Mammifères non volants | Non présents |
Iguanes et lézards | Sur-représentés |
Amphibiens | Non présents |
Orchidées (plantes à fleurs en général) | Sous-représentées |
Fougères | Sur-représentées |
La colonisation par des plantes s'envisage très bien avec le transport par le vent (pour les espèces anémophiles), l'eau (espèces hydrophiles) ou les oiseaux (espèces zoophiles). Le maintien ou non des populations colonisatrices selon leur type de reproduction est facilement compréhensible quand on compare des orchidées, hautement dépendantes de leurs pollinisateurs, avec des fougères dont les semences sont résistantes et légères.
Par contre la colonisation par des animaux aussi particuliers que les iguanes qui, mis à part les iguanes marins, ne nagent pas et qui n'ont aucune chance de profiter d'un voyage aérien nécessite de solides arguments. Le premier d'entre eux est qu'il faut considérer l'étendue de la durée pendant laquelle ces animaux ont eu la possibilité de coloniser ces iles ; c'est-à-dire depuis leur émergence il y a au moins 10 Ma pour les plus anciennes. Si la probabilité d'un unique événement de colonisation à un instant donné est quasi nulle, celle qu'une colonisation arrive au moins une fois en 10 Ma est déjà beaucoup plus grande. D'autre part, des cas similaires de colonisation insulaire ont été étudiés dans le monde (par exemple, la colonisation de Madagascar par des rats et souris aujourd'hui endémiques) et font ressortir trois hypothèses majeures :
- l'existence dans le passé de « ponts émergés », comme les ponts continentaux évoqués du temps de Wegener ;
- l'existence d'anciennes iles plus proches de la côte qui auraient servi de relais à la colonisation des iles les plus éloignées ;
- le transport d'espèces animales et végétales sur des radeaux de fortunes à la suite de violentes tempêtes continentales.
Les deux premières hypothèses peuvent être écartées d'emblée car, comme nous l'avons vu précédemment, la géologie et la géographie de cette partie du Pacifique ne vont pas dans leur sens. La dernière hypothèse, bien que farfelue de prime abord, est soutenue par plusieurs observations :
- en 1911, une ile végétale de 30 m² avec des arbres de près de 9 m de haut et des animaux a dérivé sur plusieurs centaines de kilomètres dans l'Atlantique du Nord ;
- de tels radeaux ont été vus dériver sur l'Amazone suite à de violentes tempêtes et inondations ;
- les courants marins et les vents sont tels qu'un radeau dérivant depuis la côté Sud-américaine aurait de grande chance d'atteindre les Galapagos en deux semaines à peine.
Un tel modèle s'accorde bien avec les observations faites sur la faune des Galapagos : seuls des animaux assez résistants face au manque d'eau et de nourriture survivraient à un tel voyage. Alors que des grenouilles se dessécheraient très rapidement et que des rats et souris mourraient de soif, des iguanes pourraient eux en sortir vivant et accoster sur une plage. En prenant, de plus, en compte le temps imparti, il est tout à fait probable qu'une seule expédition d'infortune de quelques couples d'iguanes, voire d'une unique femelle gravide, ait pu aboutir.

Source - © 2017 Google Earth, modifié
Figure 42. Courants marins superficiels baignant les Galapagos dans le Pacifique Est
La situation géographique des Galapagos permet aux iles d'être baignées par des courants marins de surface convergents. Ainsi, il n'est pas inenvisageable qu'un radeau puisse atteindre "rapidement" les iles à la seule faveur des courants marins. Le terme de « courant El Niño », tout comme le phénomène El Niño, est lié à sa saisonnalité ; ce courant fonctionne principalement pendant la période de Noël, or, en espagnol, El Niño désigne « l'enfant Jésus ».
Des hypothèses sur l'origine des iguanes : une ou plusieurs phases de colonisation ?
Sachant que le processus de colonisation est possible, il nous faut expliquer la présence de 4 espèces différentes d'iguanes, et surtout de deux genres aux modes de vies très différents. En 1961, sur la base de caractères morphologiques, une première hypothèse impliquant une évolution indépendante de ces deux genres a été proposée (Eibl-Eibesfeldt, 1961 [L2]).
Deux phases de colonisation
Eibl-Eibesfeldt proposa que les iguanes marins et les iguanes terrestres ont divergé indépendamment de leurs ancêtres continentaux, alors supposés ressembler à l'iguane vert (Iguana iguana).
 Source - © 2008 Paul Asman et Jill Lenoble - CC BY 2.0 | 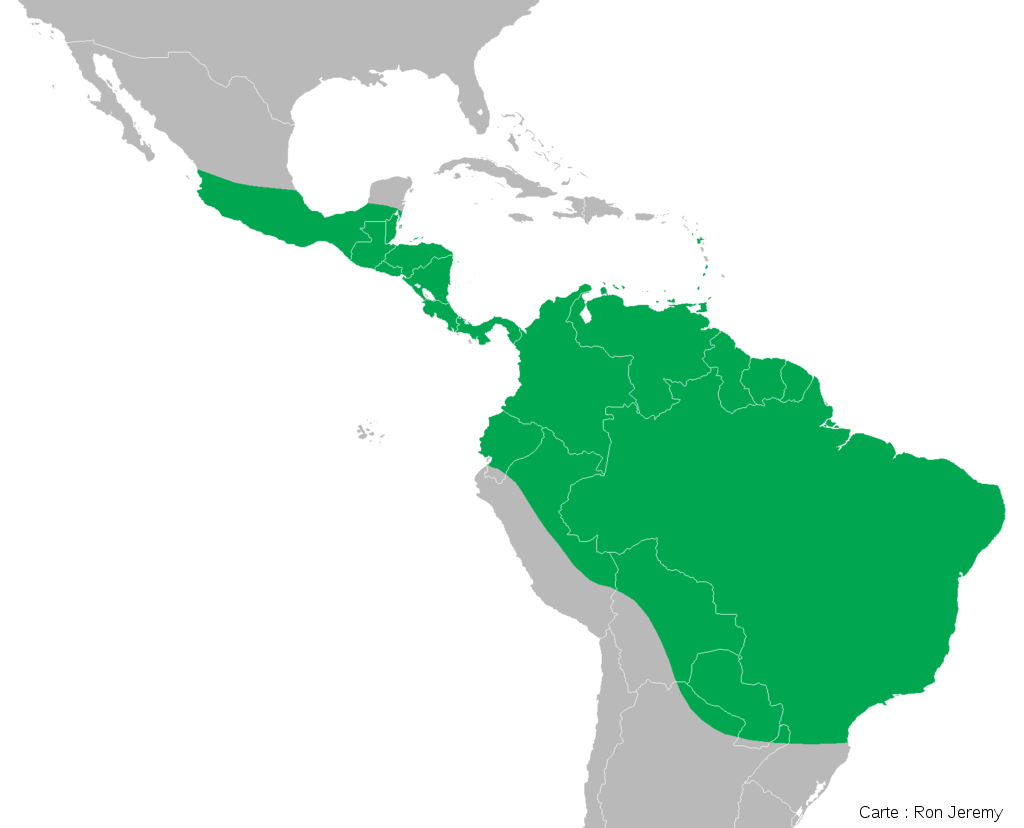 Source - © 2009 Ron Jeremy - CC BY 3.0 Figure 44. Aire de répartition actuelle de l'iguane vert en Amérique centrale et en Amérique du Sud La répartition de l'iguane vert est compatible avec l'hypothèse d'un départ en radeau depuis les côtes Sud-américaines. |
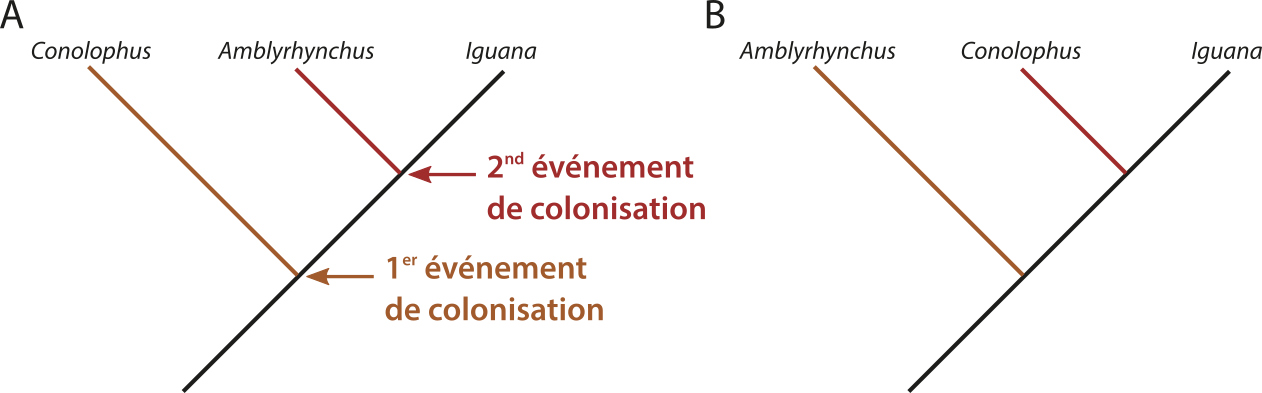 Source - © 2017 Florent Figon - CC BY-SA 3.0 Le scénario proposant l'arrivée en deux temps des iguanes aux Galapagos se traduit par deux histoires évolutives possibles. La première (A) aurait vu la divergence des iguanes terrestres des Galapagos à partir d'une population d'Iguana. Ensuite, une autre population d'Iguana serait arrivée et aurait donné naissance aux iguanes marins. À l'inverse, on peut imaginer que les iguanes marins aient divergé en premier, puis les iguanes terrestres ensuite (B). Quel que soit l'ordre de divergence, chaque genre d'iguanes des Galapagos est plus proche du genre continental Iguana que de l'autre genre des Galapagos. Cela se traduirait alors par l'un des deux arbres phylogénétiques, A ou B. | |
Ce modèle peut s'expliquer de deux façons. D'un côté, les iguanes marins et terrestres des Galapagos auraient évolué indépendamment sur le continent depuis deux populations d'iguanes verts. Ils auraient ensuite colonisé en deux temps les Galapagos. Seules les populations insulaires auraient alors survécu. Problème : aucun fossile ressemblant à des iguanes des Galapagos n'a été retrouvé en Amérique du Sud.
L'autre solution, proposée par Wyles et Sarich en 1983 [24], est que des iguanes verts ont vogué jusqu'aux iles Galapagos au début du fonctionnement du point chaud. Puisque les iles étaient récentes, la végétation était peu développée et donc défavorable aux iguanes. Par hasard ou par nécessité, quelques iguanes ont commencé à se nourrir d'algues, d'abord dans la zone intertidale. Certains auraient ensuite réussi à survivre en allant chercher des algues plus en profondeur, leur permettant de laisser plus de descendants que leurs confrères terrestres, et ainsi de suite. Le temps passant et les iles vieillissant, la végétation s'est développée et une nouvelle colonisation par des iguanes verts a comblé une niche écologique vide, celle des gros lézards terrestres herbivores. Les deux espèces endémiques des Galapagos ont alors continué à diverger sur l'ile. Wyles et Sarich (1983 [24]), sur la base de données immunologiques, proposent une date de divergence autour de 15-20 Ma (date qui pose question puisque les iles Galapagos semblent n'être âgées que de 4 à 5 Ma pour les plus anciennes voire 10 Ma pour les des iles aujourd'hui immergées, problème de datation géologique ou biologique ?).
Une unique phase de colonisation
Un autre modèle, développé en même temps que le précédent, propose que la colonisation ne se soit faite qu'en une seule fois. Nous ne traiterons que de l'hypothèse où il y a eu spéciation et divergence sur les Galapagos car ce même scénario sur le continent puis colonisation en même temps par les deux espèces parait hautement improbable.
Indépendamment du nombre de colonisations, nous ferons l'hypothèse que l'espèce sœur des iguanes des Galapagos fait partie du genre Ctenosaura qui était un autre prétendant à ce titre dans les modèles de spéciation.
 Source - © 2006 Jackhynes | 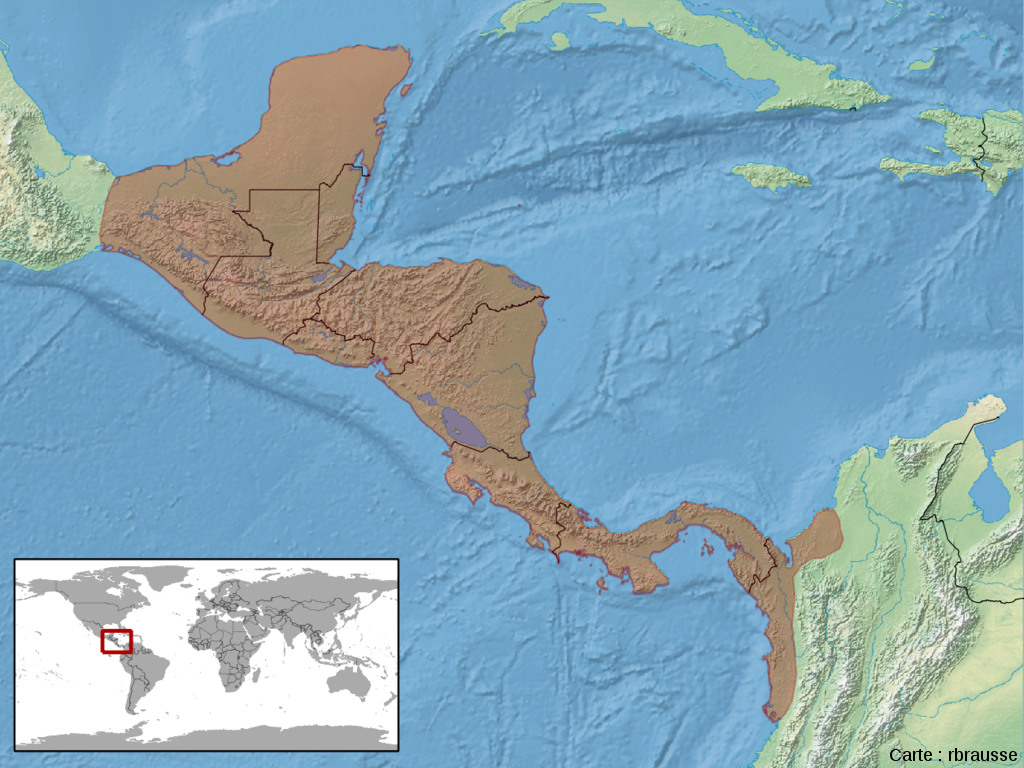 Source - © 2008 rbrausse - CC BY-SA 3.0 Figure 47. Aire de répartition actuelle du genre Ctenosaura en Amérique centrale et en Amérique du Sud De même que l'iguane vert, cet iguane aurait très bien pu traverser l'océan Pacifique vers les Galapagos depuis l'une des régions qu'il occupe actuellement. |
Dans ce modèle, une unique population d'iguanes du genre Ctenosaura aurait colonisé les iles Galapagos. Le fait que ces iguanes aient été uniquement terrestres et qu'ils aient donné des espèces encore inféodées à ce milieu (le genre Conolophus) implique qu'il y avait des ressources exploitables sur l'ile où ils sont arrivés, autres que les algues. Cela signifie que la partie de l'ile où ils ont accosté devait être suffisamment âgée et humide (donc vraisemblablement au Sud) pour qu'il y ait une végétation développée. À partir de là, le manque de ressources végétales et la compétition possible avec d'autres herbivores (comme les tortues des Galapagos) et d'autres insectivores (comme les lézards des Galapagos) ont pu être le moteur d'une différenciation comportementale, puis physiologique et morphologique, de deux populations d'iguanes qui ont chacune donné les deux genres actuels, terrestre et marin.
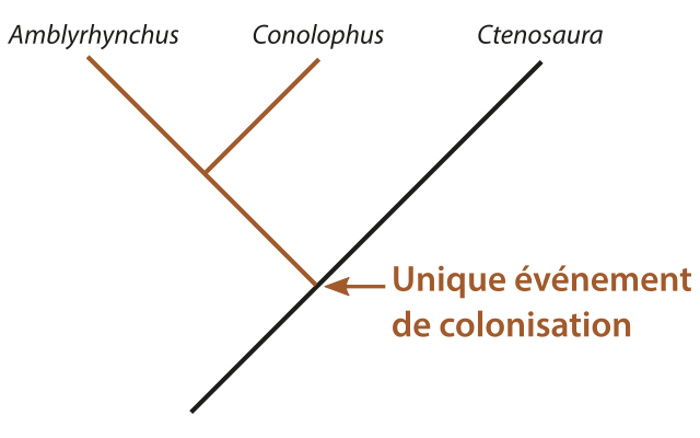
Source - © 2017 Florent Figon - CC BY-SA 3.0
Dans le cas d'un unique événement de colonisation, il n'y a qu'une seule histoire évolutive possible qui est représentée par cet arbre phylogénétique. Puisque les deux genres d'iguanes des Galapagos ont divergé depuis une même population d'iguanes continentaux (proche du genre Ctenosaura), ils sont plus apparentés entre eux qu'avec n'importe quel autre genre d'iguanes.
Faits et observations
Avant de regarder en détail les faits et observations accumulées depuis plus de 30 ans, faisons un point sur les différentes hypothèses et prédictions de chaque modèle.
Plusieurs événements de colonisation | Unique événement de colonisation | |
|---|---|---|
Parenté entre les iguanes marins | Amblyrhynchus et Conolophus ne sont pas des groupes frères | Amblyrhynchus et Conolophus sont des groupes frères |
Ancêtre commun | Iguana ou Ctenosaura | Iguana ou Ctenosaura |
Lieu de spéciation | Les iles Galapagos et/ou le continent | Les iles Galapagos |
Nombre de spéciations | Deux spéciations | Une seule spéciation |
Type d'iles colonisé en premier | Iles âgées et/ou récentes | Iles âgées |
Fossiles des deux genres | Sur le continent et/ou les iles Galapagos | Sur les iles Galapagos uniquement |
Si on considère uniquement le critère de parcimonie, le modèle avec un seul événement de colonisation devrait être privilégié. La comparaison avec d'autres espèces endémiques à faible dispersion nuance cependant cette première conclusion :
- espèces issues d'une unique phase de colonisation : les tortues des Galapagos il y a au moins 3,2 Ma (Parent et al. 2008, Poulakakis et al. 2012) ;
- espèces issues de plusieurs phases de colonisation : les geckos des Galapagos avec une première colonisation il y 13 Ma (âge un peu plus ancien que l'âge estimé des iles aujourd'hui immergées et étudiées) et une seconde il y a 3 Ma (Torres-Carvajal et al., 2014 [19]), et les lézards des Galapagos avec deux événements contemporains de colonisation sur deux iles différentes, autour de 2-3 Ma (Benavides et al., 2009 [2]).
La paléontologie nous apporte peu de preuves en faveur de l'une ou l'autre de ces hypothèses ; la plupart des fossiles trouvés aux Galapagos sont trop récents ou pas assez équivoques pour être assignés à un des deux genres (Rassman, 1997 [16]).
Les éléments les plus intéressants sont à regarder du côté des arbres phylogénétiques basés sur des critères moléculaires. Si ces études ont soulevé de nombreux doutes et critiques (cf. la « Guerre des Iguanes » due à l'effet d'attraction des longues branches en phylogénie moléculaire ; Wiens et Hollingworth, 2000 [21]), il est dorénavant bien établi que les iguanes les plus apparentés aux iguanes des Galapagos sont du genre Ctenosaura (MacLeod et al., 2015 [10]).
Là où les études phylogénétiques peuvent vraiment permettre de départager les deux modèles de colonisation, c'est en regardant les liens de parenté entre Conolophus et Amblyrhynchus. Et les résultats sont éloquents : ces deux genres sont plus proches l'un de l'autre qu'avec le genre Ctenosaura ! Des cas d'individus hybrides entre Amblyrhynchus et Conolophus (sans qu'on sache s'ils étaient fertiles) ont même été plusieurs fois observés (Rassmann et al., 1997 [17], MacLeod et al., 2015 [10]). Cela souligne leur proximité génétique et confirme donc leur proche parenté.
Des méthodes de datation évolutive (horloge moléculaire) appliquées à cette hypothèse ont longtemps suggéré un âge de divergence autour de 10 Ma (Rassmann, 1997 [16]), soit peu après l'émergence de la plus ancienne ile observée (maintenant sous-marine). Une étude récente propose un âge autour de 4,5 Ma (MacLeod et al., 2015 [10]) ce qui coïnciderait avec l'émergence des actuelles iles les plus anciennes (Española et San Cristobal). À cette époque, il devait donc y avoir des iles plus âgées possédant une végétation suffisante qui ont permis le maintien et l'évolution d'iguanes terrestres.
Conclusion : une histoire évolutive qui se complète au fur et à mesure
L'ensemble des résultats précédents privilégie l'hypothèse d'une unique phase de colonisation par des iguanes terrestres apparentés au genre Ctenosaura. La divergence entre Conolophus et Amblyrhynchus se serait alors produite sur une ile déjà âgée, mais à une époque à laquelle San Cristobal et Española étaient probablement déjà émergées.
Ce modèle n'exclut pas de possibles événements d'arrivées d'iguanes continentaux par la suite, mais cela le complexifie en ne faisant que retarder le moment de la divergence (par des flux génétiques depuis le continent vers les iles).
Si nous avons parlé des événements les plus précoces de l'évolution des iguanes des Galapagos, de nombreuses études se penchent sur les migrations et les spéciations / différenciations à l'intérieur des deux genres. Auparavant datée à plus de 5,7 Ma (Gentile et al., 2009 [8]), l'apparition de l'espèce basale du genre Conolophus (l'iguane rose, C. marthae) a été revisitée à 1,5 Ma (MacLeod et al., 2015 [10]) ; un résultat qui est en accord avec le modèle décrit dans cet article. Des problématiques telles que l'absence de spéciation dans le genre Amblyrhynchus, mais la présence de nombreuses sous-espèces récemment différenciées, sont largement discutées. L'importance de la géologie dans l'évolution des espèces est envisagée dans la récente divergence des deux autres espèces d'iguanes terrestres et la différenciation génétique des sous-espèces d'iguanes marins, toutes deux estimées autour de 0,05 Ma (cf. explication sur le conundrum (énigme) dans MacLeod et al., 2015 [10]).
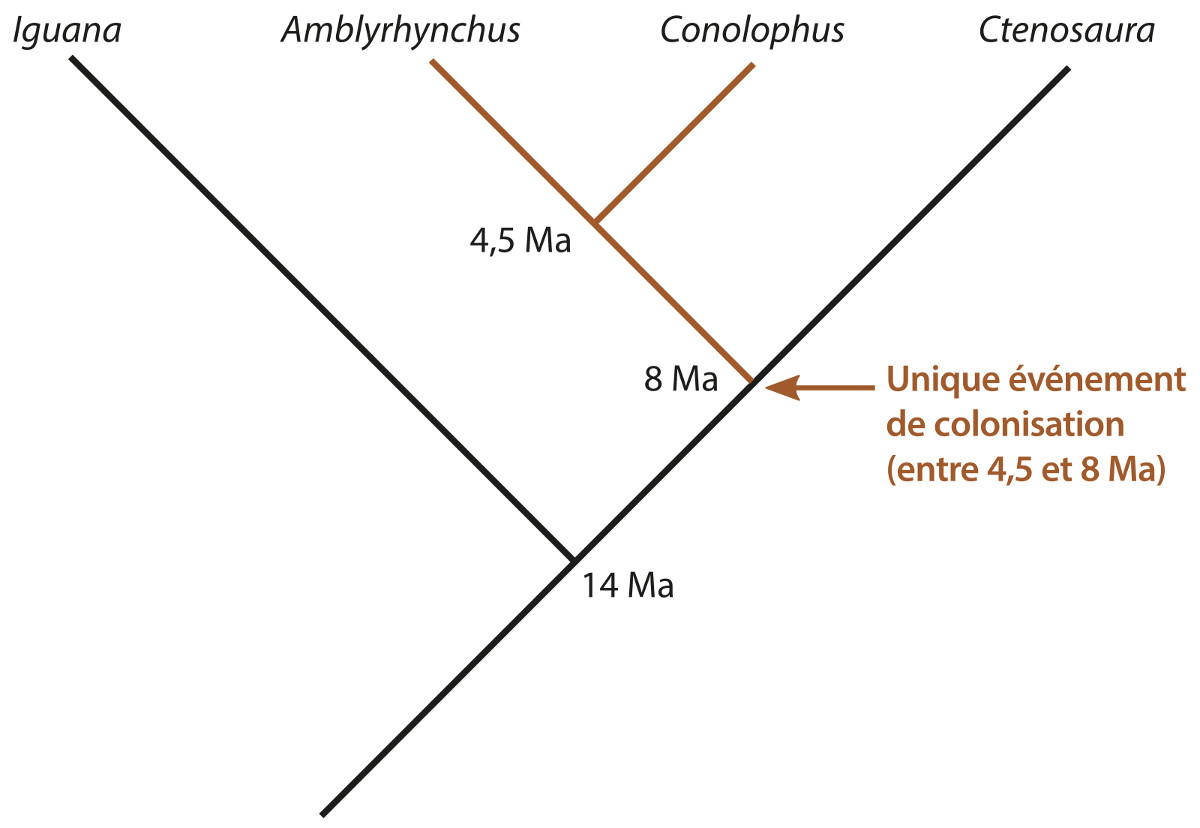
Source - © 2017 Florent Figon - CC BY-SA 3.0
Les données les plus récentes de la phylogénie moléculaire ont permis d'obtenir l'arbre évolutif ci-dessus. Ces données suggèrent que le genre continental le plus apparenté aux iguanes des Galapagos est Ctenosaura, ils auraient divergé il y a près de 8 Ma. De plus, les iguanes marins et terrestres des Galapagos auraient divergé il y a 4,5 Ma. On sait alors que l'événement de colonisation s'est produit au plus vers 8 Ma, mais forcément avant 4,5 Ma. Le clade Ctenosaura et iguanes des Galapagos se serait quant à lui séparé du genre Iguana il y a 14 Ma. Ces datations par la méthode de l'horloge moléculaire sont tout à fait cohérentes avec les âges des iles émergées et immergées.
Quelles mesures de conservations pour les iguanes des Galapagos ?
La conservation et la protection des écosystèmes sont des problématiques majeures aux Galapagos. Ces iles sont depuis 1936 un immense parc national ; il est géré par le Parc National des Galapagos et représente 97% de la surface émergée de l'archipel. L'écosystème marin fait, lui, partie de la réserve marine des Galapagos depuis 1986. L'archipel des Galapagos est inscrit au Patrimoine mondial de l'UNESCO depuis 1978.
Avec l'aide la Fondation Charles Darwin, créée en 1959 (100 ans après la publication de l'œuvre majeure de Darwin), ces entités promeuvent la protection des écosystèmes uniques de ces iles, tout en permettant une croissance contrôlée des habitations, un accès raisonné aux touristes et le travail des scientifiques.
Le statut de conservation des iguanes des Galapagos
La protection des iguanes des Galapagos se heurte à plusieurs problèmes : une structure génétique encore mal comprise (en particulier chez les iguanes marins, puisqu'ils sont à la fois les représentants d'une unique espèce et présents sur toutes les iles), une aire de distribution fragmentée pour chaque sous-espèce ou espèce, et un statut de conservation variable pour chacune d'entre elles. La consultation du site de l'UICN rend compte de la complexité du statut de conservation qu'il faut appliquer pour protéger convenablement ces populations uniques.
Espèce / sous-espèce | Aire de répartition | Statut de conservation | Historique du statut de conservation |
|---|---|---|---|
Iguane terrestre C. subcristatus | Toutes les iles, absent uniquement sur l'ile San Cristobal ? | Vulnérable | "Rare" jusqu'en 1994, "vulnérable" depuis 1996 Pas de mise à jour depuis |
Iguane terrestre C. pallidus | Ile Santa Fé | Vulnérable | "Vulnérable" depuis 1986.Pas de mise à jour depuis |
Iguane terrestre C. marthae | Volcan Wolf sur l'ile Isabela | En danger critique d'extinction | Première et unique évaluation en 2012 |
Iguane marin A. cristatus | Toutes les iles, avec une répartition insulaire pour chaque sous-espèce | Quatre sous-espèces vulnérables, deux en danger d'extinction et cinq non-répertoriées | Pas de mise à jour depuis 2004 |
Les statuts actuels de l'UICN, bien que peu mis à jour, montrent que les iguanes des Galapagos sont aujourd'hui en danger, ou a minima menacés et vulnérables. Pour mieux les préserver, il nous faut donc comprendre les dangers auxquelles ils peuvent être soumis.
Les menaces environnementales
Si les Galapagos sont connues pour avoir aidé Darwin à développer sa théorie de l'évolution par la sélection naturelle (cf. Sélection naturelle, VIH, pinsons : ce que Darwin ne pouvait pas savoir), elles ont aussi permis à un autre grand naturaliste, Wallace, contemporain de Darwin et co-découvreur de la sélection naturelle (cf. L'autre découvreur de la sélection naturelle : Alfred R. Wallace), d'exprimer ses craintes à propos de l'extinction des espèces, en particulier face à la menace de l'homme. Alfred Russel Wallace expliquait ainsi comment les perturbations provoquées par l'homme pouvait affecter et détruire des populations animales insulaires, tel que l'iguane marin des Galapagos (Kutschera et Kleinhans, 2013 [9]).
Le phénomène El Niño
Ce phénomène climatique, d'une récurrence d'environ 7 ans, est bien connu des pêcheurs péruviens. Pendant la situation "normale", en dehors des phénomène El Niño ou La Niña, le déplacement des eaux chaudes de surface vers l'Ouest du Pacifique provoque une remontée des eaux froides (upwelling) sur les côtes péruviennes. Cet apport de nutriments provoque une augmentation du nombre de poissons, donc une pêche plus favorable. Lors d'un phénomène El Niño, les eaux chaudes superficielles normalement cantonnées à l'Ouest se dirigent vers l'Est, envahissent les côtes Sud-américaines, y provoquent précipitations et inondations, et limitent le développement des poissons en bloquant l'upwelling côtier.
Mais ce phénomène El Niño n'a pas d'effets négatifs que sur le continent car le déplacement des eaux chaudes, et donc des zones de précipitations équatoriales, modifie rapidement le climat des Galapagos. Sur Santa Cruz, il peut y pleuvoir dix fois plus que la normale. Cette brusque augmentation des précipitations, mais aussi de la température des eaux, de la modification des courants marins, etc, modifie les écosystèmes et leur fonctionnement (Vinueza et al., 2006 [20]).
Les effets d'El Niño sur les populations d'iguanes marins sont multiples :
- un remplacement des algues rouges et vertes par des algues brunes, non comestibles pour les iguanes, à cause du réchauffement des eaux de surface et la baisse de l'apport en nutriments par les eaux froides (Vinueza et al., 2006 [20]) ;
- une mortalité au sein des populations d'iguanes marins pouvant atteindre 90% (Vinueza et al., 2006 [20]) ;
- une régression de la taille des individus au cours de ces épisodes, les anglo-saxons parlent de shrinking : des individus déjà adultes peuvent réduire leur taille en modifiant non seulement leur cartilage, mais aussi leurs os en les déminéralisant (Wikelski et Thom, 2000 [22]).

Source - © 2015 Florent Figon - CC BY-SA 3.0
Figure 50. Exemple d'iguanes amaigris sur l'île San Cristobal
Des individus dans une telle condition d'amaigrissement peuvent être vus aux Galapagos soit à la suite d'un événement climatique tel qu'El Niño, mais aussi lors d'épisodes plus locaux d'appauvrissement en nourriture ou lorsque des mâles et des jeunes se retrouvent isolés du reste des iguanes sur une petite ile.
Si les populations d'iguanes marins ont toujours connu ces épisodes climatiques extrêmes, leur amplitude et leurs effets ont augmenté ces dernières années. Il devient donc de plus en plus difficile pour une population de revenir à un niveau normal.
Contrairement aux iguanes marins, les iguanes terrestres sont supposés profiter des événements El Niño (l'augmentation des précipitations favorisant le développement des végétaux terrestres). C'est alors l'événement inverse, La Niña, avec une sécheresse plus importante que la normale, qui pourrait menacer les populations. Il manque encore des études et des données sur cet effet potentiel.
Le volcanisme et la faune endémique locale
Si le volcanisme de grande intensité (comme les trapps) a un rôle bien connu dans la modification de la biodiversité (cf. Le volcanisme paroxysmal et ses interactions avec la biosphère), on sait aussi que le volcanisme local, surtout quand il est effusif et non pas explosif, a peu d'impact majeur sur la biodiversité, sauf si celle-ci est inféodée au volcan en éruption et qu'elle vit dans une très petite zone sur ses flancs.
Ce type de menace a été abondamment mis en avant par les médias lors de l'éruption de 2009 du volcan La Cumbre sur l'ile Fernandina. Celle-ci menaçait alors une population endémique de tortues terrestres de ce volcan. Le même type de raisonnement est applicable à l'espèce C. marthae dont la centaine d'individus ne vit que sur les flancs du volcan Wolf de l'ile Isabela. Celui-ci étant en éruption depuis mai 2015, cette espèce reste en danger. De plus, celle-ci revêt une importance évolutive et phylogénétique particulière car elle est l'espèce « basale » du genre Conolophus ; elle est donc la plus à même de nous donner des informations sur l'ancêtre commun de tous les iguanes terrestres, qui lui-même a divergé de l'espèce d'iguane marin.
Les effets des menaces anthropiques
Aujourd'hui, on considère que l'élément le plus important dans l'évolution, mais aussi l'extinction, des iguanes est l'activité anthropique : introduction de prédateurs tels que les rats et les chiens, augmentation du stress des individus, modification du système immunitaire et endocrinien, modification de l'habitat côtier, etc.
Une cause avérée menaçant fortement des iguanes marins est l'épanchement d'huile et de gasoil suite au chavirage de bateaux. On se souvient de l'importance de celui du tanker Jessica en janvier 2001 qui coula au large de l'ile San Cristobal. Les courants marins ont emporté la pollution plus à l'Ouest vers Santa Fé ce qui a provoqué la mort de 62% de la population d'iguanes marins de l'ile (Wikelski et al. 2002). On comprend alors que les effets coup sur coup d'un tel épanchement d'huile et d'un El Niño peuvent mener la population d'une ile à disparaître.
Des stratégies de conservation
Pour établir une stratégie de conservation d'une espèce, il faut réfléchir à au moins trois questions.
- L'espèce, ou la population, en question est-elle actuellement en danger ? Ce qui est manifestement le cas pour les iguanes des Galapagos, bien qu'il faille mettre à jour les statuts de l'UICN qui commencent à dater.
- Pourquoi faudrait-il investir du temps et de l'argent dans la protection de cette espèce plutôt qu'une autre ? Dans le cas de C. marthae, l'importance phylogénétique et évolutive est un élément de réponse. L'endémisme et la particularité des iguanes marins en est une autre.
- Sur quelles menaces pouvons-nous agir pour réduire sa probabilité d'extinction ? En d'autres termes, quelles menaces pouvons-nous véritablement contrôler ?
Cette dernière question est la plus importante dans un programme de conservation. Les spécialistes proposent de se centrer sur les menaces pour lesquelles nous sommes la cause. Celles sur lesquelles nous pouvons agir rapidement, efficacement et durablement sont donc :
- la gestion d'espèces introduites, comme le chien, le chat, les rats... qui s'attaquent soit aux œufs (rats), soit aux jeunes iguanes (chats et chiens), et contre lesquels les iguanes n'ont pas de systèmes de défense (car ils n'ont quasiment pas de prédateurs terrestres sur ces iles, mis à part la buse des Galapagos ; Berger et al., 2007 [3]) ;
- les épanchement d'huile issus du fonctionnement des bateaux et du tourisme ;
- la pression touristique ;
- et, enfin, l'aménagement du territoire.
En jouant sur tous ces leviers, nous devrions pouvoir faire disparaître les pressions anthropiques qui s'ajoutent à celles qui sont environnementales et naturelles (El Niño et volcanisme). Pour des dernières, soit nous ne pouvons pas agir sur le court terme (changement global et augmentation des fréquences et de l'intensité des événements El Niño), soit les iguanes ont pu s'adapter puisqu'ils les rencontrent depuis les débuts de leur arrivée sur l'archipel (par exemple, diminution de la taille par shrinking des adultes lors d'un évènement El Niño). Dans le cas de la néo-prédation par des espèces introduites par l'homme, des adaptations comportementales apparaissent (fuite face à un animal étranger), mais leur réponse n'est pas assez forte pour leur permettre d'y échapper (Berger et al., 2007 [3]).
Ainsi des éléments de sauvegarde des iguanes des Galapagos existent et nous montrent qu'il n'est pas vain de vouloir sauver ces animaux aussi emblématiques que fascinants d'un point de vue évolutif.
Bibliographie et sitographie
Livres et thèses
C. Darwin, 1875. Voyage d'un naturaliste autour du monde, La Découverte / Poches (2006), 574p., ISBN: 978-2707151018 (autres éditions disponibles)
I. Eibl-Eibesfeldt, 1961. Galapagos: the Noah's ark of the Pacific,
J. Fitter, D., Fitter, D. Hosking, 2016. Wildlife of the Galápagos, Princeton University Press, 272p., ISBN: 978-0691170428
M.H. Jackson, 1993. Galápagos: A Natural History, University of Calgary Press (version revue et augmentée, 2016, 336p., ISBN: 978-1895176407
A.R. Wallace, 1880. Island Life, The Phenomena and Causes of Insular Faunas and Floras, Macmillan, versions en ligne via le Projet Gutenberg
Sites internet
Pages sur la dératisation sur le site Galápagos Conservancy [consulté le 23/10/2017]
Articles scientifiques
W.T. Aguilera, J. Málaga, J.P. Gibbs, 2015. Conservation: Giant tortoises hatch on Galápagos island, Nature, 517, 271–271
S. Berger, M. Wikelski, L.M. Romero, E.K.V. Kalko, T. Rödl, 2007. Behavioral and physiological adjustments to new predators in an endemic island species, the Galápagos marine iguana, Hormones and Behavior, 52, 653–663
I. Eibl-Eibesfeldt, 1962. Neue Unterarten der Meerechse, Amblyrhynchus cristatus, nebst weiteren Angaben zur Biologie der Art, Senckenbergiana Biologica, 43, 177-199
I. Eibl-Eibesfeldt, 1966. Marine iguanas, Animals, 9, 150-153
D.J. Geist, A.R. McBirney, R.A. Duncan, 1986. Geology and petrogenesis of lavas from San Cristobal Island, Galápagos Archipelago, Geological Society of America Bulletin, 97, 555
D.J. Geist, H. Snell, H. Snell, C. Goddard, M.D. Kurz, 2014. A Paleogeographic Model of the Galápagos Islands and Biogeographical and Evolutionary Implications, in The Galapagos: A Natural Laboratory for the Earth Sciences, K.S. Harpp, E. Mittelstaedt, N. d' Ozouville, D.W. Graham (éditeurs), coll. AGU Geophysical Monograph Series, 145–166 [pdf]
G. Gentile, A. Fabiani, C. Marquez, H.L. Snell, H.M. Snell, W. Tapia, V. Sbordoni, 2009. An overlooked pink species of land iguana in the Galápagos, Proceedings of the National Academy of Sciences, 106, 507–511 [pdf]
U. Kutschera, S. Kleinhans, 2013. Alfred Russel Wallace and the destruction of island life: the Iguana tragedy, Theory in Biosciences, 132, 259–265
A. MacLeod, A. Rodríguez, M. Vences, P. Orozco-terWengel, C. García, F. Trillmich, G. Gentile, A. Caccone, G. Quezada, S. Steinfartz, 2015. Hybridization masks speciation in the evolutionary history of the Galápagos marine iguana, Proceedings of the Royal Society- B: Biological Sciences, 282, 20150425 [pdf]
A. Miralles, A. Macleod, A. Rodríguez , A. Ibáñez, G. Jiménez-Uzcategui, G. Quezada, M. Vences, S. Steinfartz, 2017. Shedding light on the Imps of Darkness: an integrative taxonomic revision of the Galápagos marine iguanas (genus Amblyrhynchus), Zoological Journal of the Linnean Society
G. Merlen, 2014. Plate Tectonics, Evolution, and the Survival of Species: A Modern Day Hotspot, in The Galapagos: A Natural Laboratory for the Earth Sciences, K.S. Harpp, E. Mittelstaedt, N. d' Ozouville, D.W. Graham (éditeurs), coll. AGU Geophysical Monograph Series, 119–144
H. Nicholls, 2013. Invasive species: The 18-km2 rat trap, Nature 497, 306–308 [pdf]
C.E. Parent, A. Caccone, K. Petren, 2008. Colonization and diversification of Galápagos terrestrial fauna: a phylogenetic and biogeographical synthesis, Philosophical Transactions of the Royal Society- B: Biological Sciences, 363, 3347–3361 [pdf]
N. Poulakakis, M. Russello, D. Geist, A. Caccone, 2012. Unravelling the peculiarities of island life: vicariance, dispersal and the diversification of the extinct and extant giant Galápagos tortoises, Molecular Ecology, 21, 160–173
K. Rassmann, 1997. Evolutionary Age of the Galápagos Iguanas Predates the Age of the Present Galápagos Islands, Molecular Phylogenetics and Evolution, 7, 158–172
K. Rassmann, F. Trillmich, D. Tautz, 1997. Hybridization between the Galápagos land and marine iguana ( Canolophus subcristatus and Amblyrhynchus cristatus ) on Plaza Sur, Journal of Zoology, 242, 729–739
S. Steinfartz, A. MacLeod, 2016. The conservation status of the Galápagos marine iguanas, Amblyrhynchus cristatus: a molecular perspective, Amphibia-Reptilia, 37, 91–109
O. Torres-Carvajal, C.W. Barnes, M.J. Pozo-Andrade, W. Tapia, G. Nicholls, 2014. Older than the islands: origin and diversification of Galápagos leaf-toed geckos (Phyllodactylidae: Phyllodactylus ) by multiple colonizations, Journal of Biogeography, 41, 1883–1894
L.R. Vinueza, G.M. Branch, M.L. Branch, R.H. Bustamante, 2006. Top-down herbivory and bottom-up El Niño effects on Galápagos rocky-shore communities, Ecological Monographs, 76, 111–131
J. Wiens, B.D. Hollingsworth, 2000. War of the Iguanas: Conflicting Molecular and Morphological Phylogenies and Long-Branch Attraction in Iguanid Lizards, Systematic Biology, 49, 143–159 [pdf]
M. Wikelski, C. Thom, 2000. Marine iguanas shrink to survive El Niño, Nature, 403, 37–38
M. Wikelski, V. Wong, B. Chevalier, N. Rattenborg, H.L. Snell, 2002. Galápagos Islands: Marine iguanas die from trace oil pollution, Nature, 417, 607–608
J.S. Wyles, V.M. Sarich, 1983. Are the Galápagos iguanas older than the Galápagos? Molecular evolution and colonization models for the archipelago, in Patterns of Evolution in Galápagos Organisms, R.I. Bowman, M. Berson, and A.E. Levinton (éditeurs),, AAAS, Pacific Division, San Francisco, pp. 177–185 [pdf du livre]