Article | 03/02/2005
Homo floresiensis : un homininé nain contemporain d'Homo sapiens
03/02/2005
Résumé
Découverte d'un nouveau représentant du rameau humain dans l'île de Florès (Indonésie).
Table des matières
La description des fossiles d'un nouveau représentant du rameau humain vient tout juste d'être publiée dans la revue Nature du 28 octobre 2004[1][2]. Ces fossiles ont été mis au jour sur l'île de Florès, l'une des îles de la Sonde, dans l'archipel indonésien, et sont attribués à une nouvelle espèce, H. floresiensis. La datation des sédiments et du charbon de bois trouvé en association avec les fossiles indique que ces individus existaient depuis plus de 38.000 ans, et que leur disparition ne remonte au plus qu'à 18.000 ans. Ils sont donc contemporains de notre espèce Homo sapiens.
Cependant, ce nouveau parent présente des caractères anatomiques très spectaculaires, à commencer par une taille ne dépassant pas 1 mètre pour l'adulte, et un volume du cerveau corrélativement réduit, du même ordre de grandeur que celui d'australopithèques beaucoup plus anciens.
Les auteurs de cette découverte rattachent cette espèce au groupe de l'Homo erectus, le premier Homininé à avoir occupé l'Asie. Ils proposent que ses caractéristiques singulières soient le résultat d'une évolution de type "évolution insulaire", dont on a montré qu'elle se traduit souvent, chez les grands mammifères, par une réduction de taille. Ces fossiles indiqueraient aussi que notre genre Homo peut présenter une variabilité morpho-anatomique plus importante qu'on ne le supposait jusqu'alors.
Ces découvertes illustrent à nouveau le caractère buissonnant et non-linéaire de l'évolution humaine, et la coexistence, dans le passé, de plusieurs espèces humaines. Elles soulignent enfin, si l'on retient hypothèse de l'évolution insulaire, combien les modalités d'évolution de notre groupe sont semblables à celles des autres mammifères.
Homo floresiensis, un nouveau cousin
Son aspect
Les fossiles découverts sont le crâne et des éléments post-crâniens d'un individu adulte, ainsi qu'une prémolaire d'un autre spécimen trouvée dans des dépôts plus âgés. Le squelette principal est dénommé LB1. Brown et al. affirment qu'il « combine une mosaïque de traits primitifs, uniques, et dérivés qu'on ne rencontre chez aucun autre Homininé », et que « ses proportions dentaire et faciale, son anatomie post-crânienne cohérente avec une bipédie obligatoire de type humaine et un appareil masticateur similaire, par sa taille relative et sa fonction, à celui des humains modernes, plaident tous pour son assignation au genre Homo »[1]. Le trait le plus marquant est la taille très réduite du crâne (figure 1 en haut) comparativement à celle de l'Homo sapiens actuel (~1.500 cm3, figure 1 en bas), de Homo neanderthalensis (1.200 à 1.800 cm3) ou d'Homo erectus (780 à 1.200 cm3) : seulement 380 cm3, soit une capacité crânienne semblable aux plus faibles valeurs rencontrés chez les australopithèques (380 à 500 cm3) et proche de celle des chimpanzés actuels (350 à 400 cm3)(1).
(1) Capacités crâniennes d'après Crubézy É., Braga J. et Larrouy G., Anthropobiologie. Masson, 2002.
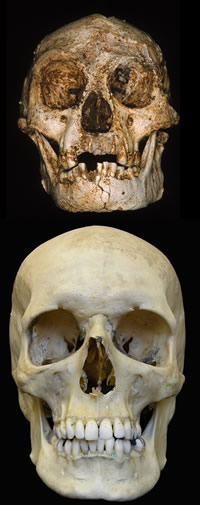
Source - © 2004 P.Brown, Nature
Le volume du crâne de l'Homo sapiens actuel est de 1.500 cm3 environ.
Le lieu
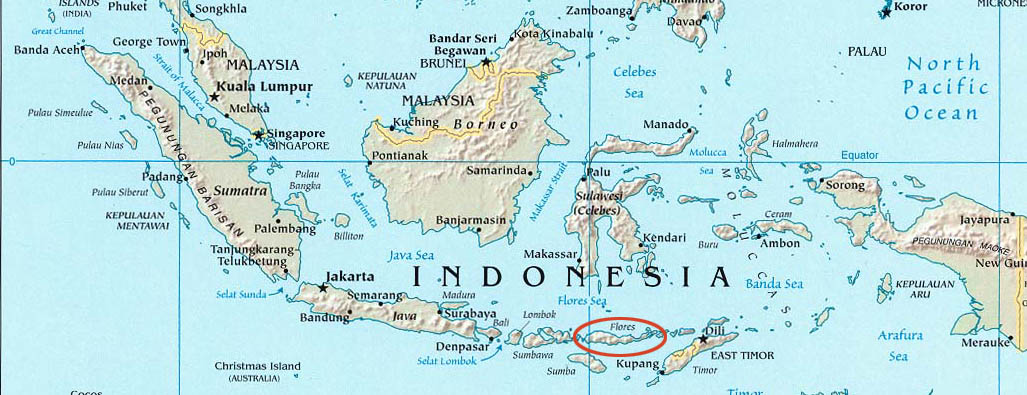
Ces fossiles proviennent de l'île de Flores (14.000 km2), qui appartient à l'archipel volcanique des îles de la Sonde (figure 2), en Indonésie(2). Cette position géographique aux confins de l'Asie suggère une évolution endémique de cet homininé à partir d'ancêtres issus du continent eurasiatique.
(2) : Pour comparaison, la superficie de la Corse est de 8.000 km2.
Ses relations phylogénétiques
L'analyse de la forme du crâne découvert, quasiment pas déformé, rattache ce spécimen à Homo erectus, dont les spécimens considérés comme les plus caractéristiques ont été justement découverts en Asie (Chine) et en Indonésie (Java), et dont la répartition temporelle s'étale de -1,5 Ma au moins à -250.000 ans, ce qui prolongerait donc encore la durée d'existence de cette branche asiatique du genre Homo, sortie d'Afrique depuis -1,8 Ma. Cette proposition de parenté implique aussi que les ancêtres de ces individus étaient de plus grande taille qu'eux, et suggère donc une évolution vers le nanisme.
Les implications de cette découverte
Un nouvelle feuille au buisson des Homininés
L'évolution humaine ne peut plus être vue comme linéaire, si l'on considère toutes les espèces d'Hominidés actuellement reconnues, et même si l'on peut discuter la validité de toutes les espèces proposées, par exemple pour le seul genre Homo :H. erectus, H. ergaster, H. heidelbergensis, H. georgicus, H. sondaicus, H. sapiens…et donc H. floresiensis).
Dans le cas de H. floresiensis, s'il est apparenté à H. erectus, et vu ses particularités morphologiques, il est plus que probable qu'il ait constitué une espèce écologique et biologique bien distincte de son contemporain H. sapiens, dont il était a priori séparé depuis plus d'un million d'années. Avec H. neanderthalensis, le Pléistocène a donc compté au minimum trois espèces du genre Homo plus ou moins contemporaines, qui ont pu, théoriquement, se côtoyer (H. sapiens et H.neanderthalensis au Moyen-Orient et en Europe jusque vers -30.000 ans, H. sapiens et H.floresiensis en Indonésie jusqu'à -18.000 ans).
Cependant, H.floresiensis repose également le problème de l'ampleur réelle de la variabilité morphologique au sein du genre Homo. Avec sa petite taille et ses caractères particuliers, ce fossile « montre que le genre Homo est morphologiquement plus varié et flexible dans ses réponses adaptatives que l'on ne le pensait jusqu'à présent » selon les termes de P. Brown et al., ce qui pourrait donc aussi remettre en question la validité de certaines espèces de notre genre.
Un cas d'évolution insulaire ?
Pour P. Brown et ses collègues [1], il ne s'agit pas d'une réduction de taille similaire à celle des populations humaines actuelles de petites tailles comme les pygmées africains. En effet, expliquent-ils, chez ces populations « l'explication de la réduction de taille fait appel à des avantages en termes de thermorégulation pour une vie en milieu chaud et humide, soit par un refroidissement par évaporation accru (augmentation du rapport surface/volume), soit par un taux de production de chaleur interne réduit. Chez les pygmées africains, la stature réduite est le résultat d'une diminution des taux de facteur de croissance IGF-1 (insuline-like growth factor 1) au cours de la période de croissance, ou d'une réceptivité diminuée à l'IGF-1. Bien que la taille adulte soit réduite, les proportions crânio-faciales reste dans la gamme des populations adjacentes, de taille adulte plus importante, de même que la taille du cerveau. La combinaison de la petite taille et de la taille du cerveau chez LB1 n'est pas cohérente avec un retard de croissance post-natal lié à l'IGF-1. De même, ni le nanisme pituitaire (ou nanisme par déficit isolé en hormone de croissance), ni le nanisme microcéphale primordial des humains modernes ne reproduit les traits squelettiques présents chez LB1 ».
Ses découvreurs suggèrent donc que le nanisme de H. floresiensis serait le résultat d'une évolution endémique, liée à un isolement de longue durée, et serait un cas d'évolution insulaire(3) vers le nanisme assez comparable à celle documentée pour d'autres animaux de grande taille, comme les mammouths ou les cervidés [3].
On connaît en effet des spécimens fossiles de mammouths et d'éléphants nains (figure 3), retrouvés dans des zones (à l'époque) isolées, coupées du continent et d'aire restreinte, respectivement la Sicile et la péninsule de Wrangel, en Sibérie orientale.
Des cas de nanisme insulaire ont également été documentés pour d'autres grands mammifères, comme des cerfs (figure 4) et des chèvres.
(3) Le qualificatif d'insulaire n'implique pas forcément une évolution sur une île au sens géographique, mais dans une région relativement isolée, géographiquement et donc génétiquement, de la population initiale.

Source - © 2000 La valse des espèces, Dossier Hors Série, Pour la Science
Il mesure 70 centimètres de haut. Ce sont les crânes fossiles de cet animal qui ont probablement inspiré la légende du Cyclope d'Ulysse.
L'interprétation souvent proposée pour cette tendance évolutive est celle d'une sélection en faveur des petites tailles (besoins alimentaires diminués) et d'un temps de gestation raccourci (plus de temps disponibles pour la recherche de nourriture) dans des environnements aux ressources alimentaires restreintes. (Pour les grands mammifères comme l'éléphant, on peut mentionner aussi la diminution de la pression de sélection exercée par les prédateurs, contre lesquels il est avantageux d'être de grande taille).
Il faut noter qu'à l'inverse, l'évolution insulaire de mammifères initialement de petite taille (en gros inférieure à celle d'un lapin), comme les rongeurs, se traduit souvent par l'acquisition de tailles plus importantes, voire gigantesques (figure 4). Dans ces cas-là, une explication avancée serait le relâchement de la pression de sélection en faveur d'une petite taille, pour deux raisons : d'abord la diminution du nombre de compétiteurs pour les ressources alimentaires - en milieu insulaire, le nombre de concurrents de taille similaire diminuerait, permettant aux petits animaux présents d'élargir leur niche écologique, d'exploiter des ressources ordinairement utilisées par des animaux plus gros - la niche écologique effectivement réalisée de ces animaux se rapprocherait ainsi de leur niche potentielle ; ensuite la raréfaction des prédateurs, contre lesquels une taille réduite peut être un avantage pour un animal déjà petit.
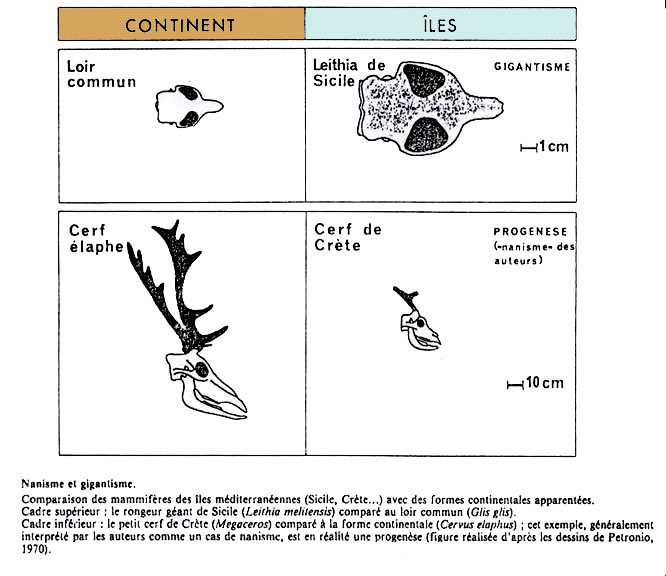
Source - © 1986 J.-L. Dommergues et al., Geobios n°19, fasc.3
Cadre supérieur : le rongeur géant de Sicile (Leithia melitensis) comparé au loir commun (Glis glis).
Cadre inférieur : le petit cerf de Crète (Megaceros) comparé à la forme continentale (Cervus elaphus). Cet exemple, généralement interprété par les auteurs comme un cas de nanisme, est en réalité une progenèse (figure réalisée d'après les dessins de Petronio, 1970).
Cependant, il est impossible, en l'absence d'autres fossiles, de savoir si H. floresiensis a bien acquis cette taille réduite sur place, postérieurement à son arrivée sur cette île, ou s'il descend d'une population d'Hominines de taille déjà réduite.
L'évolution humaine, une évolution comme les autres
Cette interprétation de ce nouveau fossile bat évidemment en brèche l'idée d'une tendance évolutive vers un cerveau de plus en plus volumineux, ou celle d'une meilleure adaptation des formes possédant le plus gros cerveau.
Elle confirme également que différentes espèces du genre Homo (et plus généralement différents représentants des Homininés) ont coexisté, au moins temporellement, et que Homo sapiens est donc aujourd'hui le dernier représentant d'un genre autrefois plus varié (à l'instar du cheval actuel).
Si, de plus, cet Hominine est effectivement le produit d'une évolution endémique insulaire, il confirme combien l'évolution du rameau humain, et du genre Homo, a suivi jusqu'à tout récemment les mêmes processus que ceux rencontrés chez d'autres mammifères de taille similaire.
Bibliographie
[1] Brown P., Sutikna T., Morwood M. J., Soejono R. P. et al. (2004). A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431(7012): 1055-1061
[2] Morwood M. J., Soejono R. P., Roberts R. G., Sutikna T. et al. (2004). Archaeology and age of a new hominin from Flores in eastern Indonesia. Nature 431(7012): 1087-1091
[3] Hartenberger J-L., La mort des mammifères des îles. Pour la Science 276, Octobre 2000
Sur le site de l'INRP :
Origine et évolution de l'homme
Sur le site Planet-Vie, la biologie au lycée :