
Article | 15/09/2010
De Burgess à Franceville (Gabon) : les plus anciennes traces fossiles de pluricellulaires
15/09/2010
Résumé
Du Cambrien au Paléoprotérozoïque, à la recherche des plus vieux fossiles d'êtres pluricellulaires (Burgess, Chengjiang, Édiacara, Doushantuo, Stirling, Franceville).
Table des matières
Introduction
Au début de l'été 2010, la publication par la revue Nature d'un article signé d'une équipe internationale dirigée par le chercheur français Abderazak El Albani [25] a secoué le monde de la paléontologie en affirmant que l'apparition des premières structures multicellulaires organisées remonteraient à 2,1 milliards d'années, soit 200 à 500 millions d'années plus tôt que l'on ne l'envisageait jusqu'à présent.
Le commentaire de cette découverte et des questions qu'elle soulève est aussi l'occasion de faire le point sur les plus anciennes communautés de macrofossiles connues à ce jour, à commencer par les plus célèbres, celles de Burgess et d'Édiacara.
Vu l'abondante littérature scientifique publiée à propos de ces gisements très anciens, fouillés pour certains depuis des années, cet article ne peut prétendre à l'exhaustivité. Il tente de se concentrer sur des données relativement récentes.
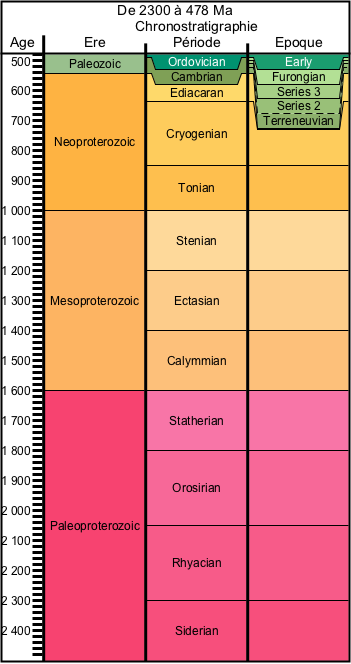
La figure 1 permettra de replacer dans le temps les assemblages fossiles présentés. On notera aussi la rareté des fossiles disponibles sur cette gigantesque portion de l'histoire de la Terre qu'est le Protérozoïque.
Burgess, Chengjiang et « l'explosion cambrienne »
Burgess Pass, Canada
Le site de Burgess Pass, dans les schistes des Rocheuses canadiennes, est probablement l'un des gisements fossilifères les plus connus aujourd'hui. D'une part par son contenu et son âge, estimé à 505 Ma, d'autre part par l'histoire de sa découverte et de l'analyse de ses fossiles. Datant du Cambrien inférieur et montrant de nombreux fossiles d'animaux macroscopiques, il constitua longtemps, depuis sa découverte en 1909 par le paléontologue Charles Doolitle Walcott (1850–1927), la seule « fenêtre » disponible sur la biocénose de de cette période particulière — « l'explosion cambrienne » — où les macrofossiles paraissaient faire brutalement leur apparition.
Source - © 2009 ISC
Mais sa réputation tient aussi beaucoup à la ré-interprétation de ses fossiles et à l'essai qu'en tira le célèbre paléontologue américain Stephen Jay Gould en 1989.
⇒⇒
Petit résumé de l'essai de S. J. Gould. - Dans son ouvrage Wonderful Life (en français, "La vie est belle"), Gould explique comment Walcott, sous l'influence de ses propres conceptions et des idées de son temps, avait intégré « au chausse-pied » tous les organismes décrits à Burgess parmi les grands plans d'organisation des organismes actuels (arthropodes, mollusques, etc.). Il décrit ensuite comment le ré-examen de ces fossiles dans les années 1970, par Harry Witthington, Derek Briggs et Simon Conway Morris, a conduit à y reconnaître effectivement des organismes appartenant aux grands plans d'organisation existant encore aujourd'hui ou connus dans le registre fossile plus récent, mais échouait à classer dans ces plans d'organisation connus un certain nombre d'autres fossiles. Stephen Jay Gould développait ensuite l'hypothèse iconoclaste selon laquelle l'explosion cambrienne aurait été marqué par une floraison non seulement d'organismes, mais surtout de plans d'organisation différents, dont seuls quelques-uns auraient persisté par la suite.
Le Cambrien aurait ainsi été une période pendant laquelle les processus du développement ontogénétique des organismes étaient plus « souples » et auraient permis à la vie métazoaire « d'explorer » un plus grand nombre de solutions structurales qu'aujourd'hui, créant ainsi une plus grande disparité que l'actuelle, la disparité désignant l'amplitude des différences morpho-anatomiques entre plans d'organisation, à distinguer de la diversité, qui mesure le nombre d'espèces au sein d'un même clade : la disparité entre un mollusque, un vertébré et un insecte est ainsi bien plus grande que celle qui sépare des dizaines d'espèces de coléoptères les unes des autres.
Les idées présentées au grand public par cet essai ont stimulé les recherches sur les fossiles cambriens et sont toujours évoquées pour tenter d'expliquer « l'explosion cambrienne » (cf. plus loin). Les résultats publiés depuis sont venus affiner et modifier l'état des connaissances que présentait S. J. Gould : certains fossiles énigmatiques ont pu, finalement, être réinterprétés et inclus dans des phyla connus (avec le cas spectaculaire d'Hallucigenia, voir ci-après) ; d'autres sont maintenant considérés comme des « branches basales » de grands clades actuels (comme Anomalocaris rattaché aux Arthropodes) ; en outre, le développement des phylogénies moléculaires et de la cladistique sont venues bouleverser les relations de parenté entre les grands clades (identification des deux ensembles Lophotrochozoaires et Ecdysozoaires, en particulier), rapprochant phylogénétiquement des clades aux plans d'organisation nettement différents.
⇐⇐
Source - © 2009 Cyril Langlois
Le site de Burgess est daté du Cambrien moyen (505 Ma). Or, le registre fossile disponible aujourd'hui indique que l'essentiel du spectaculaire accroissement de la disparité et de la diversité des métazoaires qui marque le début du Cambrien est déjà quasiment terminé à cette date. Paradoxalement, ce site constamment cité à propos de « l'explosion cambrienne » en est une image tardive.
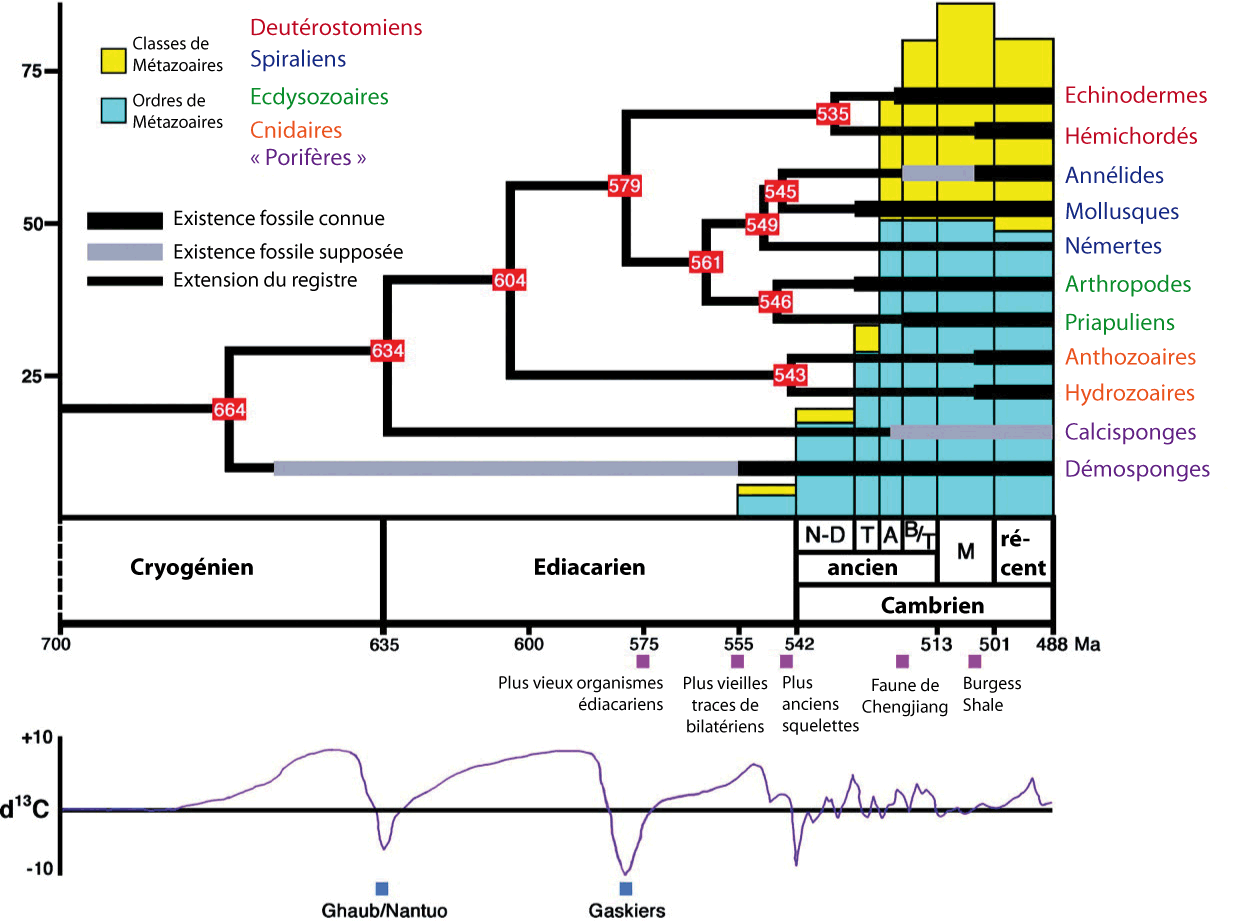
Figure 3. Tempo de l'évolution animale dans son contexte géologique à la transition Néoprotérozoïque/Cambrien
Les nœuds de l'arbre sont placés aux âges estimés par la technique de l'horloge moléculaire par Peterson et al., 2004.
Courbe isotopique du carbone d'après Knoll, 2000.
Arbre phylogénétique d'après Peterson et al., 2005 [7], francisé.
Remarque : « explosion ». . . géologique. - Il faut souligner ce que représente, en termes de durée, cet épisode qualifié d'explosion. Le nombre de familles et de genres de métazoaires fossiles passe de quelques genres à plus d'un millier en une vingtaine de millions d'années, soit le temps qui nous sépare de l'Oligocène ! Il paraît stagner ensuite (figure 3), mais si l'on élargit cette phase de diversification à l'ensemble du Cambrien et aux derniers moments du Néoprotérozoïque, sa durée est alors du même ordre de grandeur que le temps écoulé depuis la fameuse crise Crétacé-Tertiaire, il y a 65 millions d'années. Même à l'échelle géologique, cet épisode unique de l'histoire du vivant n'est pas vraiment « rapide ».
Les fossiles de Burgess sont des empreintes en deux dimensions sur des schistes, issus du métamorphisme, dans le faciès schistes verts, d'une formation marno-calcaire. Le site se trouvait au Cambrien moyen sur une marge passive, au bas d'une falaise calcaire sous-marine. Les organismes ont été brusquement enfouis sous des coulées de boue tombées de l'escarpement [5]. Il semble qu'ainsi « tous les organismes présents au moment de leur enfouissement ont été préservés indépendamment de la composition et de la fragilité de leurs tissus » [4].
Depuis la « médiatisation » des fossiles de Burgess par le livre de S. J. Gould, plusieurs fossiles alors inclassables ont été ré-analysés et sont mieux compris. La figure 4 regroupent quelques-uns des fossiles les plus connus de Burgess. On y trouve, en particulier :
- des trilobitomorphes, comme Marella, et des trilobites indubitables ;
- des chélicérates (Sanctacaris) ;
- des animaux désormais classés comme des branches basses éteintes du clade actuel des Arthropodes, comme Opabinia ou Anomalocaris, le plus grand prédateur de Burgess ;
- des crustacés, comme Canadaspis, comparable aux actuels malacostracés phyllocarides ;
- des vers priapuliens (Ottoia) et polychètes (Canadia) ;
- des onychophores (Aysheaia et probablement Hallucigenia) ;
- des mollusques, puisque la réinterprétation des curieux Wiwaxia et Odontogriphus indique qu'ils possédaient une radula [4] ;
- et bien sûr des chordés, avec Pikaia, toujours « montés en épingle » par les présentations du site de Burgess selon une tendance très nombriliste, alors que ce groupe est très minoritaire dans cette faune.

Source - © 2003 Caron et coll., Comprendre en enseigner la planète Terre, Ophrys
Figure 4. Fossiles classiques du site de Burgesss
Les avancées réalisées depuis quelques décennies dans l'analyse des fossiles de Burgess doivent beaucoup à l'éclairage apporté par d'autres sites fossilifères du Cambrien inférieur découverts depuis, dans lesquels des fossiles assez semblables à ceux de Burgess, mais mieux conservés encore, ont été retrouvés.
Le cas du fossile Hallucigenia est emblématique de ces progrès. Des fossiles chinois interprétés comme des onychophores munis de plaques épineuses dorsales ont conduit à l'idée que ce fossile au nom évocateur avait initialement été reconstitué « sens dessus dessous » : les échasses rigides qu'on lui avait supposé étaient des épines, alors que les curieux tubes qui s'alignaient sur son dos devaient correspondre à ses lobopodes marcheurs (figure 5). Il s'agirait donc d'un onychophore ou d'un groupe proche de ceux-ci (même si cette attribution reste discutée). À défaut de certitude sur leur appartenance aux onychophores, ces vers diversement « cuirassés », bien représentés dans les faunes cambriennes, sont répertoriés sous le terme de Lobopodiens.
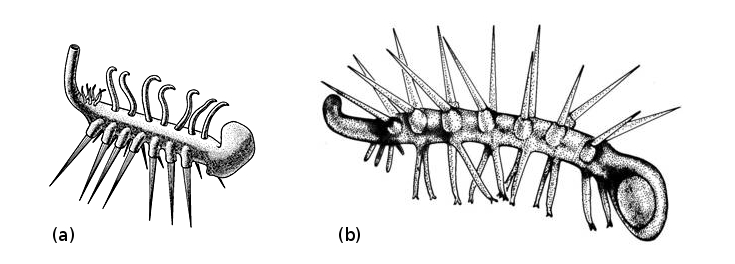
Source - © 2009 Mary Parrish, The Smithsonian Institution
Figure 5. Hallucigenia, avant et après les découvertes de Chengjiang
(a) Interprétation initiale d'Hallucigenia par S. Conway Morris.
(b) Réinterprétation d'Hallucigenia par L. Ramskold et H. Xianguang en 1991.
Parmi les progrès obtenus, citons également l'interprétation des traces fossiles (ichnologie) par l'étude d'organismes actuels en laboratoire. Une équipe franco-germano-polonaise vient ainsi de démontrer que l'ichnofossileTreptichnus pedum, des terriers horizontaux en rameaux successifs, dont la première apparition marque la limite Ediacarien- Cambrien, était très semblable aux traces de locomotion des vers priapuliens qui explorent aujourd'hui les fonds marins peu oxygénés des fjords norvégiens [2]. Ces vers priapuliens, animaux prédateurs, devaient constituer une pièce importante et pionnière dans ces nouveaux écosystèmes benthiques qui se mettaient en place au début du Cambrien.
Et après Burgess ? - Burgess est devenu le site emblématique de l'apparition des écosystèmes « modernes », mais cela ne signifie pas que les taxa qui y sont représentés vont perdurer ensuite. La majorité des grands clades (Ordres, Classes, etc.) et des plans d'organisation qu'ils représentent vont se diversifier ou se raréfier, mais resteront représentés par la suite (la disparité n'augmentera plus guère) ; par contre, de nombreux taxa cambriens de plus bas niveau taxonomique (Familles, Genres) semblaient absents des terrains ordoviciens. Cette disparition, et le grand épisode de diversification constaté à l'Ordovicien (la « Grande Diversification Ordovicienne »), suggéraient une crise à la transition Cambrien-Ordovicien. L'hypothèse concurrente étant une lacune de fossilisation, certains de ces taxa cambriens n'ayant pas de squelette minéralisé (éponges, vers, cnidaires). La présentation, au printemps 2010, de fossiles ordoviciens rappelant ceux de Burgess, et provenant d'un nouveau gisement marocain (les argiles et les silts de la formation Fezouata), vient confirmer cette hypothèse [3]. Un certain nombre d'organismes de morphologie « basale », « cambrienne » — par rapport aux fossiles typiquement ordoviciens —, rappelant Marella ou Anomalocaris, par exemple (figure 4), perduraient dans ces écosystèmes ordoviciens. La transition entre les faunes cambriennes et ordoviciennes aurait donc été plus progressive qu'on ne le croyait auparavant.
Le gisement de Burgess n'est donc plus la seule ni la plus ancienne fenêtre sur le Cambrien dont nous disposions. Un nouveau site canadien, proche de celui de Burgess, a d'ailleurs été présenté tout récemment et vient encore grossir la liste [1]. Parmi ces sites, l'un des plus remarquables reste celui de Chengjiang, en Chine méridionale.
Ichnofossile : trace d'activité biologique conservée dans les sédiments. Les ichnofossiles reçoivent un nom d'espèce, comme les organismes fossiles.
Chengjiang, Chine
Le site de Chengjiang a été mis au jour en 1984. La datation de ses roches, des schistes là encore, lui donne un âge de 525 Ma, donc du Cambrien inférieur. Il est donc antérieur à Burgess. Exceptionnel par l'état de conservation de ses fossiles, il a fourni plus de 180 espèces de métazoaires et montré que la plupart des grands groupes représentés à Burgess étaient déjà présents vingt millions d'années plus tôt [4]. Il a également affirmé l'image, donnée par Burgess, d'une faune cambrienne largement dominée par les arthropodes (figure 7) (40 %), puis les éponges (porifères, 13 %) et où les chordés, quoique présents, étaient moins représentés que les onychophores et les priapuliens, des groupe très minoritaires dans les écosystèmes actuels.

Source - © 1997 Chen, Zhou, ZU, Yeh
Figure 6. Couverture avant (à gauche) et arrière (à droite) d'un livre chinois sur le site de Chengjiang
Source : Chen, Zhou, ZU, Yeh, 1997. The Chengjiang Biota. A Unique Window of the Cambrian Explosion
Chengjiang a aussi confirmé, entre autres choses, que les chordés étaient largement plus anciens que le site de Burgess. On y a en effet trouvé non seulement des fossiles de chordés, rappelant l'amphioxus actuel ou Pikaia, mais aussi de vertébrés, avec Haikouichthys ercaicunensis. Ce fossile est interprété comme un représentant des pétromyzodontidés (les lamproies actuelles), indice que cette lignée et celle des myxines, plus éloignée encore dans l'arbre phylogénétique des vertébrés, s'étaient déjà différenciées à cette époque.
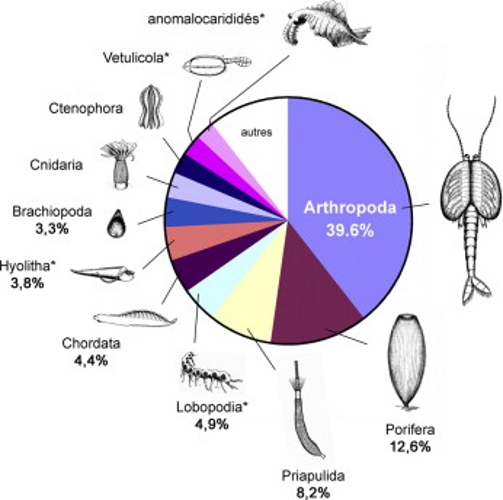
Source - © 2009 Vannier [4]
Figure 7. Biodiversité de la faune de Chengjiang
Proportions relatives des différents phylums (n = 182 espèces). (*) = groupes dont les affinités sont incertaines.
Chengjiang, Burgess et les autres gisements fossilifères du Cambrien inférieur — en Sibérie, au Groënland, en Grande-Bretagne et en Australie — démontrent que les biocénoses cambriennes comprennent majoritairement des organismes épibenthiques, mais voient aussi l'apparition des organismes fouisseurs, explorant verticalement le sédiment, ainsi que des animaux pélagiques, nageant dans la colonne d'eau, soit activement (necton et pélagos) soit passivement (plancton). Cette exploration de nouveaux secteurs du milieu marin s'accompagne d'une complexification des réseaux trophiques [4]. Les écosystèmes cambriens diffèrent ainsi radicalement de ceux qui les ont précédés, et qu'occupaient la « faune d'Ediacara ».
Les communautés fossiles de l'Ediacarien
Les sites fossilifères édiacariens
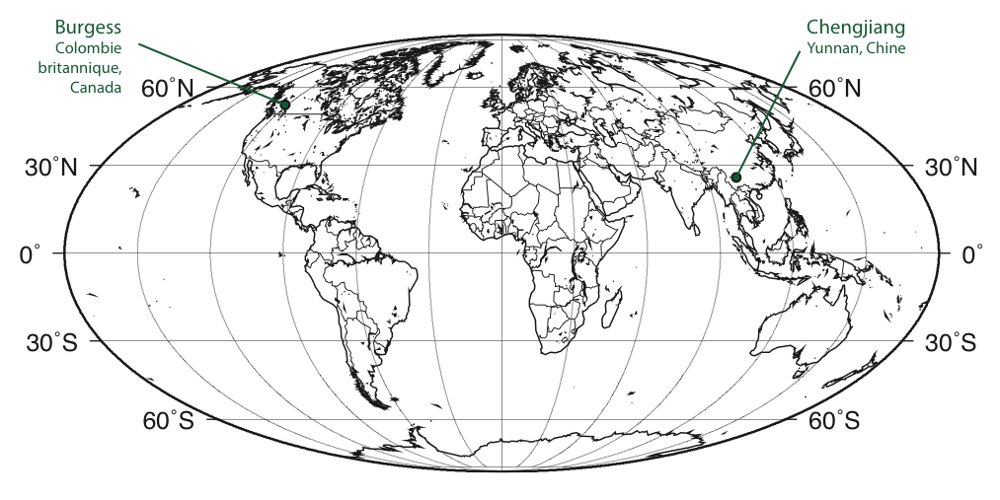
La « faune » — ou le biote — d'Ediacara désigne un ensemble de fossiles macroscopiques datant de la fin du Néoprotérozoïque, et nommé en référence au site des collines Ediacara, au Sud de l'Australie. Aujourd'hui, ces fossiles ont été identifiés en de nombreux autres endroits de par le monde (figure 8).

Source - © 2006 D'après McCall [9]
La position des principaux sites mentionnés dans le texte est précisée.
Le premier fossile d'organisme édiacarien, trouvé à Terre-Neuve, a été décrit dès 1872 par Billings sous le nom d'Aspidella terranovica, et le signalement des principaux sites fossilifères, à Terre-Neuve, à Ediacara ou encore en Namibie, remontent à la fin du XIXe et à la première moitié du XXe siècle (site namibien en 1933, Ediacara en 1946). Mais ils étaient alors considérés comme datant du Cambrien ancien, et des structures comme Aspidella ont longtemps été prises pour des traces inorganiques. C'est en 1959 que M.F. Glaessner identifia ces fossiles australiens, namibiens, canadiens ou encore anglais comme des fragments dispersés d'un même ensemble datant du Protérozoïque terminal [13].
Les âges de ces fossiles ne couvrent qu'une trentaine de millions d'années, entre 575 et 541 Ma. Ces organismes apparaissent peu de temps après la dernière phase glaciaire du Néoprotérozoïque, la glaciation Gaskiers. Celle-ci suit la glaciation marinoenne, plus intense encore, qui termine la période Cryogénienne, dernier en date des épisodes de « Terre boule de neige » (Snowball Earth).
Remarque : l'Édiacarien, nouvelle division chronostratigraphique. - Depuis mars 2004, la Commission Stratigraphique Internationale a approuvé la définition d'un nouveau couple Système / Période dans l'échelle chronostratigraphique, l'Édiacarien, correspondant au temps séparant la fin de la glaciation marinoenne du début du Cambrien, soit de -630 à -542 Ma (Science vol. 305, 30 Juillet 2004).
Quelques fossiles discoïdes très simples, plus anciens encore, ont été signalés (et datés de plus de 675 Ma). Mais, peu nombreux et mal caractérisés, ils ne sont habituellement pas considérés comme partie intégrante de cette « faune ».
À l'opposé, si les formes les plus typiques de l'Édiacarien semblent disparaître avant le Cambrien, quelques fossiles d'affinité édiacarienne ont été découverts dans des terrains cambriens, en Californie par exemple. La faune édiacarienne n'aurait donc pas entièrement disparu avant l'explosion cambrienne, mais son habitat aurait été fortement restreint avant sa disparition définitive [8].
Le biote d'Ediacara représenterait ainsi une première « bouffée de diversification » des organismes multicellulaires, rapidement avortée.
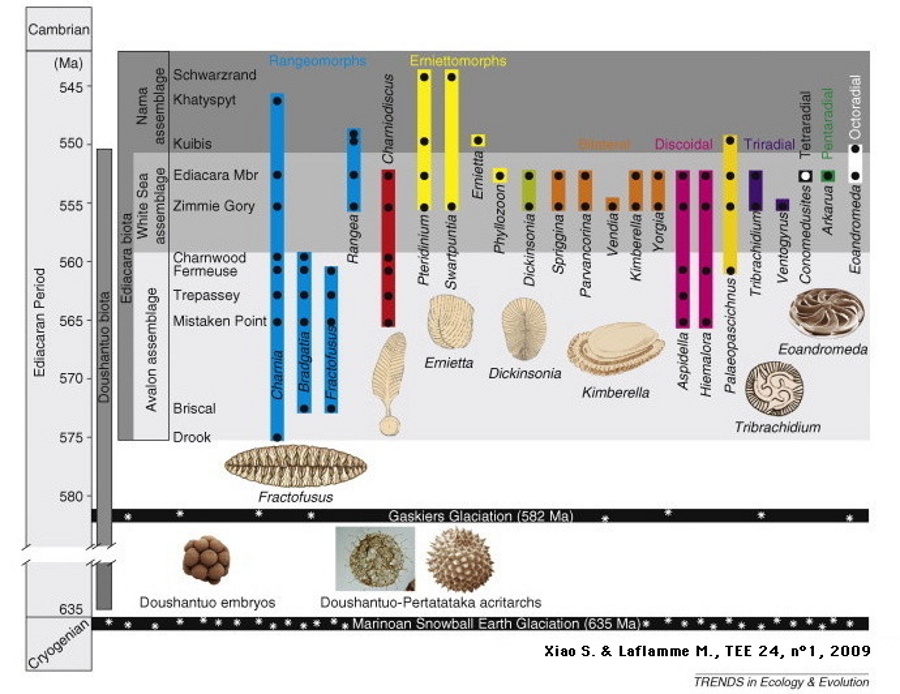
Source - © 2009 D'après Xiao & Laflamme [8]
Les trois assemblages faunistiques édiacariens sont différenciés par leur ton de gris. Les glaciations marinoenne et Gaskiers, ainsi que l'extension temporelle des fossiles de Doushantuo, sont également mentionnés.
Remarque : les Vendobiontes. Les textes relatifs aux organismes d'Ediacara utilisent parfois les termes de faune vendienne et de Vendobiontes. Que recouvrent exactement ces appellations ?
- Le nom Vendien désignait auparavant l'intervalle chronostratigraphique comprenant l'actuel Édiacarien et la fin du Cryogénien (le Varangien). Il n'est donc plus en vigueur avec la définition de l'Édiacarien.
- Le phylum des Vendobiontes est une catégorie taxonomique proposée en 1992 par Adolf Seilacher pour désigner les organismes fossiles édiacariens à morphologie en « pneu ». En 1985 puis en 1992, en effet, ce paléontologue a proposé de considérer ces fossiles comme des organismes phylogénétiquement éloignés de ceux qui se diversifient à partir du Cambrien [14], et qui constitueraient ainsi un clade à part, peut-être groupe-frère du phylum des Métazoaires. Cette idée reste controversée mais, quoi qu'il en soit, les Vendobiontes ne représentent pas l'intégralité des formes décrites dans les gisements édiacariens.
L'analyse des communautés édiacariennes
Les fossiles de l'Édiacarien sont maintenant catégorisés en trois ensembles, chacun dénommé d'après le ou les gisements sur lesquels ils sont le mieux identifiables. Ces assemblages se distinguent par leur composition (fossiles les plus caractéristiques, représentation relative des différents groupes, etc.), par leur âge et par l'environnement de dépôt des sites ([10], [8]).
- Assemblage Avalon (575-560 Ma). Fossiles de Terre-Neuve et d'Angleterre ; contexte d'eau profonde. Il s'agit de fossiles — et non de traces — préservés in situ à Terre- Neuve sous des couches de sédiments volcano-clastiques [10]. Les organismes présents sont tous fixés et dépourvus de squelette rigide. Les plus courants sont des Rangeomorphes, organismes en forme de frondes segmentées à structure fractale.
Assemblage mer Blanche (560-550 Ma). Défini à partir des sites des bords de la mer Blanche (Nord de la Russie) et d'Ediacara (Australie), il représente des milieux de plate-forme peu profonde (soumise aux vagues de tempête). Les roches de ces gisements sont principalement des quartzites à Ediacara et des grès en mer Blanche.
Des trois ensembles, c'est celui qui montre la plus grande diversité. La fossilisation, à Ediacara comme aux abords de la mer Blanche, a été facilitée par le développement d'épais tapis bactériens, qui ont préservé la morphologie des organismes sous forme d'empreintes et de moules. Les fossiles sont principalement des organismes segmentés, peut-être des animaux bilatériens, des disques, des frondes, associés à de nombreuses traces de terriers, mais là encore sans aucune forme à squelette.
- Assemblage Nama (550-541 Ma), représentés par les sites de la formation éponyme, en Namibie. Ils correspondent à des milieux agités peu profonds. Les roches fossilifères sont cette fois des calcaires. L'assemblage comprend principalement « des frondes multifoliées, des terriers d'organismes bilatériens et les premières formes connues de métazoaires à squelette calcifiés » [10].
Il semblerait donc que ces formes de vie macroscopiques se soient d'abord développées en eaux profondes, essentiellement avec le groupe des Rangéomorphes, fixés, avant de se diversifier dans des milieux plus superficiels et de s'enrichir d'espèces mobiles. Toutefois, si les assemblages mer Blanche et Nama présentent des terriers, des animaux rampants sur les fonds (épibenthiques) ou à demi-enfouis (semi-endobenthiques) et quelques formes endobenthiques, la bioturbation n'affectaient pas les couches profondes du sédiment, par ailleurs recouvert d'épais tapis bactériens (qui ont contribué à la préservation des fossiles). Les sédiments édiacariens restaient finement lités et anoxiques. Le Cambrien, avec l'apparition des animaux fouisseurs (endobenthiques), fut aussi l'occasion d'une « révolution » pour la structure des sédiments, les échanges eau-sédiments et les cycles biogéochimiques.
Les biocénoses édiacariennes différaient encore de celles qui apparaissent au Cambrien et après par cette dominance des êtres sessiles ou posés sur le sol et la rareté des prédateurs et des parasites. Cependant, le mode d'alimentation précis de ces fossiles reste souvent hypothétique. Dans le cas de l'assemblage Avalon, la profondeur des milieux exclut que ces organismes aient pu être photosynthétiques ou aient abrité des symbiotes photosynthétiques. Leur structure fractale, qui leur procurait une grande surface d'échange, suggère que les Rangéomorphes étaient peut-être des filtreurs ou des osmotrophes. En l'absence de preuves claires de déplacement et de pièces masticatrices, on ignore si des fossiles comme Parvanconina étaient des dépositivores ou des osmotrophes.
Ces communautés avaient donc une structure écologique simple, aux chaînes trophiques courtes, reposant sur une production primaire assurée par les tapis bactériens et cyanobactériens (dans la zone photique), que broutait Kimberella. On constate enfin que si la diversité de ces fossiles paraît culminer dans l'assemblage mer Blanche, la plupart des types trophiques étaient déjà présents dans les biocénoses Avalon.
Que sont les fossiles édiacariens ?
Les organismes édiacariens sont toujours énigmatiques, car les communautés édiacariennes se caractérisent par une grande disparité (de nombreux plans d'organisation différents) mais une faible diversité : on ne dénombre qu'une centaine d'espèces, mais beaucoup représentent aussi un genre complet à elle seule (remarquez que les fossiles évoqués ici sont le plus souvent désignés par leur seul nom de genre).
Plusieurs de ces espèces ont été attribuées à divers phyla connus. À ce compte, les faunes édiacariennes seraient dominées par les éponges et les cnidaires : méduses et coraux. Mais ces analyses sont largement contestées et un même organisme a pu recevoir des interprétations très diverses, allant des coraux aux colonies microbiennes ou aux unicellulaires géants.
Selon S. Xiao & M. Laflamme [8], il semble que le biote édiacarien comprenne :
- un certain nombre de fossiles et de traces encore inclassables ;
- des organismes à symétrie bilatérale ou à symétrie impaire, pour lesquels des affinités avec les phyla connus (Bilatériens et Échinodermes, en particulier) sont plausibles (cf. figure 9 les cas de Kimberella, probable mollusque [12], ou d'Arkarua, possible échinoderme) ;
- deux groupes, les Rangéomorphes et les Erniettomorphes, qui pourraient représenter des phyla originaux, totalement disparus.
Ces deux derniers groupes sont donc les plus emblématiques de la communauté édiacarienne.
- Les Erniettomorphes (Ernietta, figure 9) ont l'aspect de tubes alternant de part et d'autre d'une tige centrale. Ils sont donc dépourvus de symétrie bilatérale.
- Les Rangéomorphes (comme Charnia, figure 10) ressemblent à des frondes de fougères ou à des plumes finement découpée en barbes et barbules selon une structure fractale. Cette organisation en tubes très branchus a suggéré un rapprochement avec les coraux, mais l'organisation fine de ces fossiles ne correspond pas à celle des vrais coraux.
- La position phylogénétique de ces deux ensembles reste incertaine (figure 11), même si l'opinion qui semble prévaloir actuellement est que « les Rangéomorphes pourraient être monophylétiques et propres au biote d'Ediacara » [8].
Toutefois, ces analyses restent fragiles et sujettes à caution. Dans un commentaire publié par Science en 2004 [11], M. Brasier et J. Antcliffe soulignaient que l'on avait généralement considéré tous ces fossiles (particulièrement les Rangéomorphes) comme autant d'organismes distincts, sans prendre suffisamment en considération la possibilité que certaines des espèces décrites ne soient que des fragments d'une autre, ou des écophénotypes d'une même espèce, voire différents stades ontogénétiques d'un seul et même organisme.

Source - © 2006-2009 McCall / Xiao & Laflamme
Figure 10. Quelques fossiles édiacariens et les reconstitutions proposées pour certains d'entre eux
Charnodiscus (a) a l'aspect d'une grande fronde symétrique rattachée à une base circulaire. Fedonkin & Waggoner [12] interprètent Kimberella comme un organisme bilatérien ressemblant à un mollusque, sur la base des fossiles trouvés sur les sites de la mer Blanche (d2). Spriggina (b2) serait un organisme bilatérien segmenté mobile rappelant les trilobites ou les polychètes. Parvanconina est lui aussi rapproché des trilobites ; il s'agit peut-être d'une forme larvaire (g). Arkarua, à symétrie pentaradiée, comme Tribrachidium, triradié, sont généralement considérés comme des échinodermes. La rangéomorphe Charnia (f) se présente comme une fronde subdivisée selon une structure fractale.
Les dessins d'Arkarua adami et de Spriggina proviennent de la revue de McCall, 2006 [9]. Photos a), b1), c2), f) et g) d'après Xiao & Laflamme, 2009 [8].
La faune d'Ediacara est donc encore loin d'être entièrement comprise, ni dans sa structure écologique, ni dans la nature exacte des organismes qui la composent, pas plus que dans les causes de son développement à ce moment précis de l'histoire du Globe. Néanmoins, d'autres fossiles à peine plus anciens montrent que les métazoaires existaient bien dès la fin de la glaciation marinoenne. Ils proviennent encore d'un site chinois, les phosphorites de Doushantuo.
Les fossiles phosphatisés de Doushantuo
La formation Doushantuo, au Sud-Ouest de la Chine, dans la province de Guizhou, est une succession de calcaires et de dolomies, de schistes et de phosphorites, recouverte d'une épaisse formation carbonatée et reposant en discordance sur des tillites, marqueurs de la glaciation marinoenne [16]. Quelques passées de cendres volcaniques ont permis de dater son dépôt, par radiochronologie U-Pb, entre 635 et 551 Ma [19], soit la majeure partie de l'Ediacarien. Les lits de phosphorites, contenant les fossiles, ont des âges plus resserrés, compris entre 600 et 575 Ma environ.
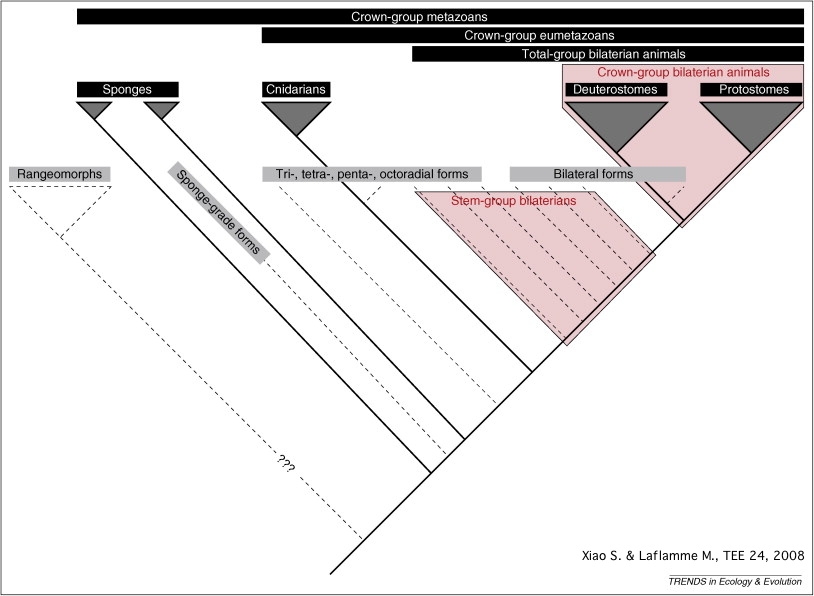
Source - © 2009 D'après Xiao & Laflamme [8]
Figure 11. Tentative de positionnement phylogénétique des principaux organismes édiacariens (en tirets)
Les Rangéomorphes formeraient un clade monophylétique, frère des Métazoaires, alors que les autres formes (à symétrie bilatérale, triradiée, pentaradiée ou octoradiée) constitueraient des branches basales de l'arbre des Métazoaires, probablement artificiellement regroupés en ensembles paraphylétiques.
Les fossiles trouvés dans ces phosphorites sont variés. Ils comprennent des restes de spongiaires, de cnidaires et de possibles bilatériens, comme les gisements édiacariens, ainsi que des petites coquilles, représentatives de la « petite faune coquillère » (small shelly fauna) retrouvée dans de très nombreux gisements du Néoprotérozoïque. Parmi ces fossiles, on peut mentionner Cloudinia, un tube calcaire conique composé de cônes emboités les uns dans les autres. Ce fossile édiacarien est fréquent mais d'affinité inconnue (et on ne l'a jusqu'ici jamais trouvé en association avec les fossiles typiques de la « faune édiacarienne »).
Mais cette région a surtout fait parler d'elle par ses fossiles de tissus mous, impeccablement préservés en trois dimensions par la phosphatisation. L'examen au microscope électronique de petites boules phosphatisées a révélé en 1998 des sphères soit creuses, soit pleines mais divisées en deux, quatre, huit ou seize et parfois entourées d'une membrane.
Tant par leur morphologie que par leur taille, ces fossiles évoquent des embryons en cours de division, figés à divers stades de développement et éventuellement encore recouverts par leur membrane pellucide (figure 12). Des résultats récents rapportent que certains de ces embryons montrent une division spirale [18], le mode de division caractéristique des protostomiens (arthropodes, annélides, mollusques, brachiopodes, etc.), un groupe probablement présent, on l'a vu, dans la faune édiacarienne. On remarque aussi que l'apparition de ces taxa à l'Ediacarien s'accorde avec certaines datations récentes des nœuds de l'arbre phylogénétique des métazoaires fondées sur le principe de l'horloge moléculaire (figure 3).
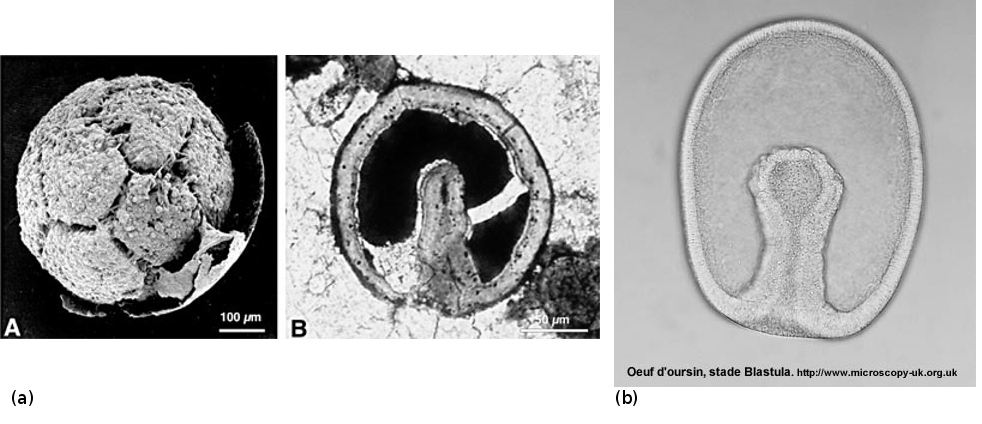
Source - © 2010
(a) Embryon phosphatisé vu au MEB (A) et coupe longitudinale d'un autre spécimen, au microscope optique (B).
(b) Embryon d'oursin actuel au stade blastula.
L'identification de ces sphérules comme des embryons a toutefois été contestée, Certains chercheurs proposent d'y voir des fossiles de bactéries « géantes », analogues à l'actuelle bactérie sulfoxydante Thiomargarita sp. (bactérie Gram négative sphérique pouvant dépasser 500 μm de diamètre). Cette hypothèse paraît toutefois invalidée par la description « d'embryons » inclus dans des structures vésiculaires ornementées, interprétées comme des oeufs enkystés en diapause, une forme de survie typiquement eucaryote [17].
En outre, d'autres fossiles d'aspect semblable ont été trouvés dans des terrains plus récents, du Cambrien inférieur et moyen. Leur auscultation à l'aide des techniques d'imagerie les plus modernes (microscopie électronique à balayage, en transmission, microtomographie aux rayons X (SRXTM)) et leur reconstitution par ordinateur [20] permet d'en visualiser les différents blastomères et jusqu'à certains organites.
phosphorite : roche sédimentaire riche en phosphate de calcium
Les fossiles méso- et paléoprotérozoïques
Les métazoaires étaient donc bien présents dès la sortie de la glaciation marinoenne vers -630 Ma. Qu'en est-il avant cette glaciation ?
Les traces du Mésoprotérozoïque
À la pointe Sud-Ouest de l'Australie, dans des roches gréseuses des monts Stirling, remontant à 1,2 Ga, des traces rappelant des terriers indiquent que des organismes pluricellulaires vermiformes et mobiles devaient déjà exister au Mésoprotérozoïque [21].
Ces traces, qui s'accompagnent d'empreintes discoïdales, sont imprimées dans un voile de boue déposée à la surface d'un chenal fluviatile, ensuite rempli de sable devenu des grès [22].
Les auteurs de cette « faune de Stirling » lui excluent une origine inorganique. Ils comparent ces marques à la trainée de mucus laissée par un gastéropode. Leur taille leur semble trop grande pour qu'elles aient été laissée par des eucaryotes unicellulaires (même si des eucaryotes unicellulaires syncitiaux actuels, certains foraminifères par exemple, atteignent des tailles macroscopiques). Elles devraient donc être l'oeuvre de multicellulaires. Toutefois, ces chercheurs ne peuvent pas exclure définitivement qu'elles aient été produites par des procaryotes coloniaux.

Source - © 2007 Bengston et al. [21]
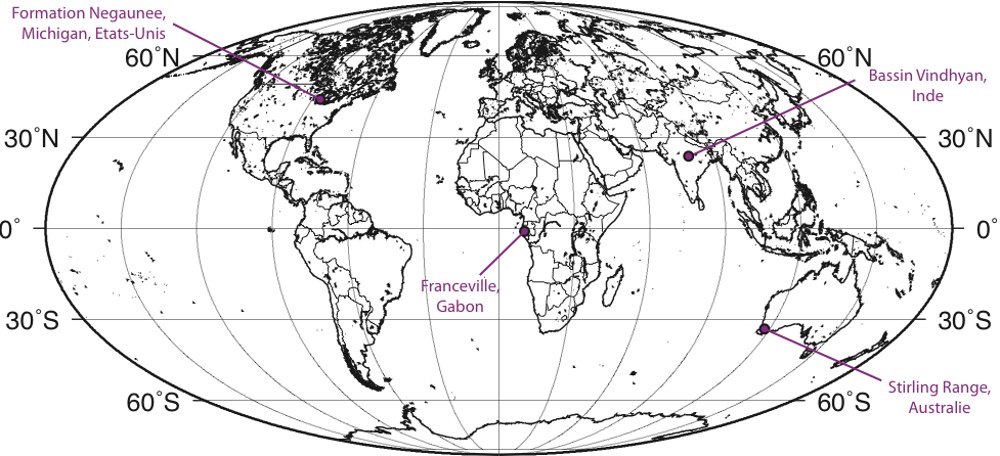
D'autres ichnofossiles très anciens ont également été signalés en Inde, dans la formation Vindhyan (figure 13) [23]. Il s'agit là aussi de sillons linéaires rappelant des terriers, à la surface de bancs de grès. Les datations radiochronologiques (K-Ar) effectuées alors donnaient des âges supérieurs à 1 milliard d'années.
Les auteurs de cette publication n'hésitaient pas à la présenter comme des indices de l'existence, dès 1 Ga, d'animaux triploblastiques (donc de bilatériens). Cette conclusion a bien sûr été mise en doute. Elle paraissait alors donner raison aux premières datations phylogénétiques obtenues par les biologistes au moyen de l'horloge moléculaire. Les résultats publiés depuis, tenant compte de la diversité de rythme des horloges et de leur possible variation de « tempo », ne supportent plus cette conclusion (figure 3 et [15]).
Source - © 2010 Cyril Langlois
Par contre, des fossiles tubulaires de plusieurs centimètres, Grypania spiralis, dont les plus vieux spécimens remonteraient à 2,2 Ga, ont été décrits dans le Michigan et le Montana, aux Etats-Unis, en Chine et en Inde. Ce fossile est attribué à une « algue » certainement eucaryote [24], mais on n'a pas de données suffisantes pour savoir si c'est une algue pluricellulaire ou une algue unicellulaire géante comme les acétabulaires actuels.

Source - © 2010 Piere Thomas / Musée des Confluences, Lyon
Ces organismes sont considérés comme des eucaryotes par la totalité des chercheurs. Il y a par contre un débat quant à leur nature pluricellulaire ou unicellulaire (cellule géante comme les acétabulaires actuels).
La possibilité qu'un organisme de ce type, bien plus simple qu'un triploblastique, ait été à l'origine des empreintes de la formation Vindhyan n'est pas définitivement exclue.
Eucaryotes syncitiaux : dont le protoplasme contient plusieurs noyaux, sans division cellulaire.
Les draperies pyritisées du Gabon
Hormis Grypania spiralis, les indices d'une vie pluricellulaire macroscopique plus organisée que les tapis bactériens ne semblait pas remonter à plus de 1,6 Ga, ou restaient très controversés. D'où l'étonnement suscité par la publication d'El Albani et al., en juillet 2010, qui repousserait de 500 Ma l'existence de tels organismes.
Ces nouveaux fossiles proviennent d'argiles noires ni métamorphisées ni déformées, situées dans le bassin de Franceville, au Sud-Est du Gabon (figure 14). Les roches étudiées appartiennent à un groupe de cinq formations superposées, en concordance : une formation gréseuse fluviatile au bas de la séquence, suivi de dépôts deltaïques où ont été découverts les fossiles, recouverts par de dépôts d'eau peu profonde puis de sédiments continentaux entrecoupés de produits volcaniques. La formation fossilifère correspond à des lits gréseux comblant des chenaux et déposés sous la limite des vagues de beau temps, dans un environnement peu agité de delta progradant. Plus de 250 fossiles ont été découverts dans des couches noires argileuses (black shales) au sein de cette formation.

Source - © 2010 F. Ossa Ossa, INSU / CNRS
Figure 16. Site fossilifère gabonais du Paléoprotérozoïque, près de Franceville, Gabon
C'est dans ces sédiments vieux de 2,1 Ga qu'ont été découverts des macrofossiles centimétriques.
La datation de ce gisement provient de l'identification d'un pic (excursion) dans l'enregistrement isotopique du carbone marin (δ13C), lui-même daté autour de 2,22 à 2,10 Ga. Les auteurs signalent également que ce dépôt s'est effectué quelques « 200 à 250 millions d'années après la première augmentation significative de la concentration atmosphérique en dioxygène atmosphérique ».

Source - © 2010 El Albani et al., Nature [25]
Les fossiles ont l'aspect de plaques ondulées pyritisées, plus ou moins allongées et parfois recouvertes d'oxydes de fer issus de l'oxydation de la pyrite. La longueur de ces fossiles va de 7 à 120 mm, la largeur de 5 à 70 mm, et l'épaisseur de 1 à 10 mm. Plusieurs d'entre elles contiennent un nodule de pyrite, formé au cours de la diagenèse précoce. Les auteurs ont reconstitué la morphologie de ces structures sur ordinateur, par microtomographie tridimensionnelle aux rayons X (SRXTM) et les ont analysé — ainsi que la roche encaissante — à l'aide d'une batterie de techniques de pointe : mesure des concentrations en sulfure et en fer, analyse de la matière organique par pyrolyse Rock-Eval et des valeurs isotopiques du carbone et du soufre par sonde ionique (spectrométrie de masse à ionisation secondaire). Ces techniques distinguent clairement les fossiles de la roche qui les emprisonne. Selon les auteurs de l'article, leurs analyses concordent pour attribuer à ces objets , qui « ne ressemblent à rien de connu », une origine biologique.
Ces fossiles évoquent des « feuillets flexibles » dont la périphérie porte souvent, mais pas systématiquement, des sillons radiaux qui paraissent « froncer » ou entailler la bordure de l'objet (figure 18). Pour ces chercheurs « [à leur connaissance,] aucun processus inorganique ne peut engendrer ce style de flexuration et la fabrique radiale irrégulière observée [. . . ] » [25]. Ce caractère suggère une structure pluricellulaire dotée d'un degré d'organisation et d'une communication intercellulaire suffisants pour produire des déformations ou des mouvements coordonnés.

Source - © 2010 INSU/CNRS
Figure 18. Exemples de spécimens et de reconstitutions par ordinateur des fossiles du Gabon
(a) Exemple de spécimen, montrant l'échelle des fossiles.
(b) Reconstruction virtuelle de la morphologie externe (à gauche) et interne (à droite) d'un spécimen.
Les auteurs reconnaissent que l'on peut cultiver aujourd'hui des bactéries capables de constituer des colonies macroscopiques et d'effectuer des mouvements chimiotactiques coordonnés, et n'excluent donc pas totalement la possibilité de colonies bactériennes (les tapis bactériens existant déjà depuis l'Archéen).
Toutefois, les structures d'origine bactérienne connues ailleurs « ne ressemblent pas aux fossiles du Gabon ». Il s'agirait donc « d'organismes coloniaux », d'assemblage de cellules communiquant entre elles, capables de déplacements coordonnés, autrement dit de pluricellulaires. El Albani et al. proposent enfin que ces organismes soient eucaryotes et non pas bactériens, en s'appuyant sur la morphologie interne de ces fossiles et sur leur analyse de la matière organique, qui a révélé la présence de stéranes, issus de la diagénèse des stérols, des molécules considérées comme propres aux eucaryotes.
Les fossiles gabonais seraient ainsi des eucaryotes pluricellulaires coloniaux, qui repousseraient l'émergence de ces formes de vie au Paléoprotérozoïque. Cela impliquerait aussi que certaines des molécules — et des gènes — permettant l'adhésion et la communication cellulaire existaient déjà à cette époque.
Cette découverte n'a pas encore suscité d'objection ni de contestation bien étayée dans les revues scientifiques. Néanmoins, certains blogs scientifiques sérieux ont commencé à lui porter un regard critique, avec quelques arguments. Ainsi Chris Nedin, auteur du blog Ediacarian. Ce blogueur signale d'abord que des figures d'échappement d'eau dans les grès d'Ediacara peuvent ressembler aux fossiles gabonais. Il compare ensuite ces derniers à divers tapis bactériens actuels, comme ceux qui se développent dans les sources hydrothermales de Yellowstone, et note que ces tapis peuvent avoir une certaine flexibilité, qu'ils ne constituent pas forcément des couches continues et qu'ils peuvent emprisonner des bulles de gaz issues des fermentations bactériennes ou de l'activité hydrothermale. Selon lui, des tapis bactériens pyritisés puis compactés par la diagenèse pourraient se craqueler radialement et produire le même genre de fossiles que ceux du Gabon.
Autrement dit, El Albani et al. rejetteraient trop vite l'hypothèse de structures produites par des tapis microbiens épais et déformés par la compaction et par des bulles de gaz (voir le billets sur les traces de 2,1 Ga et ses mises à jours (update))
Conclusion
Si l'on accepte les conclusions de l'analyse des fossiles gabonais, l'acquisition de la multicellularité suivrait de peu l'augmentation des teneurs en dioxygène des océans et de l'atmosphère, ce qui renforce l'hypothèse, souvent avancée, d'une relation causale entre ces deux événements.
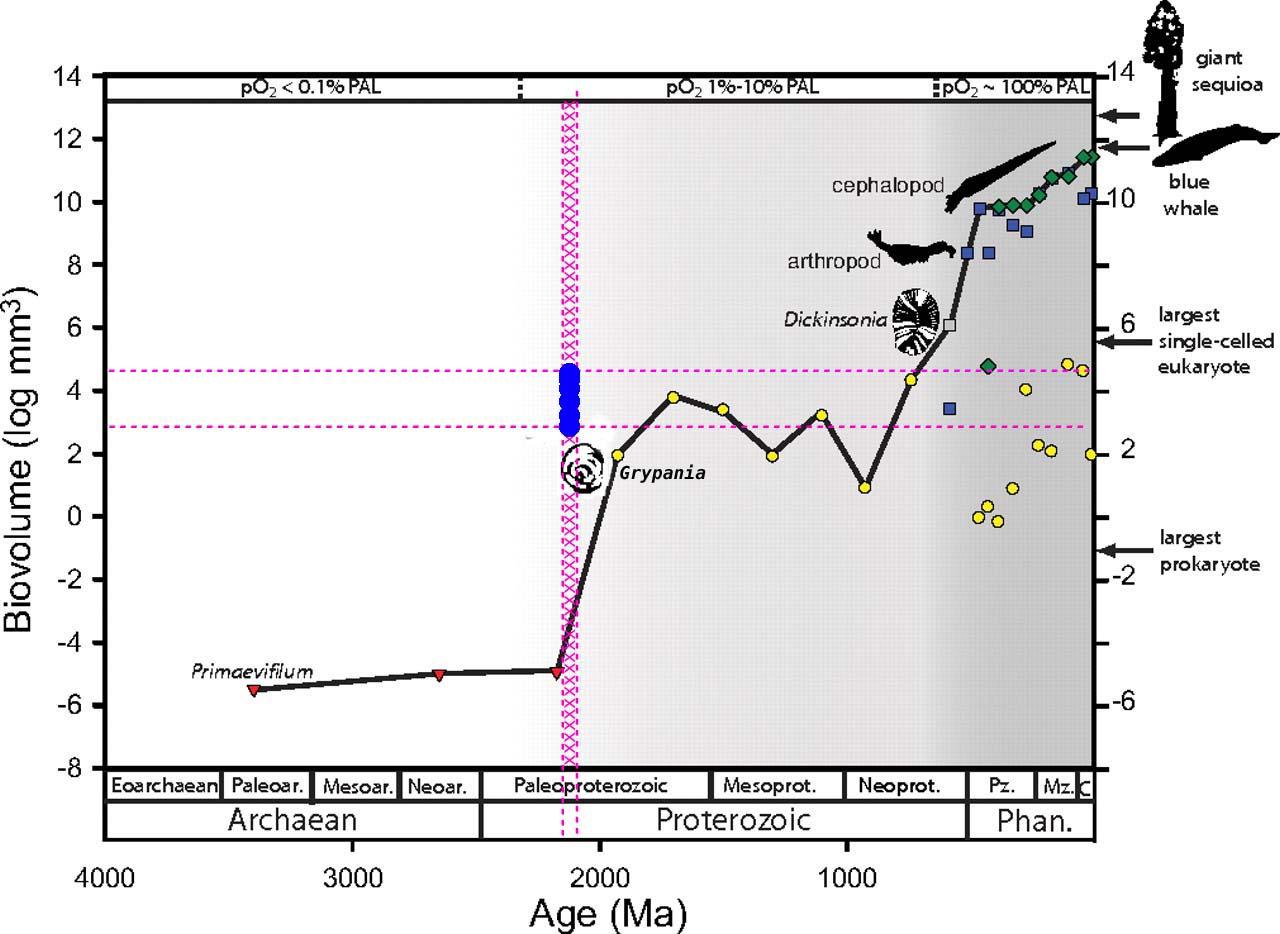
L'apparition de grands animaux pluricellulaires, tridimensionnels et non plus seulement plats, à partir de l'Édiacarien, constituerait un deuxième « saut » de taille et d'organisation pour certaines branches eucaryotes (figure 19). La période édiacarienne, quant à elle, voit le développement d'animaux pluricellulaires tridimensionnels, et non plus seulement plats, alors qu'au Cambrien se diversifient des animaux munis de systèmes spécialisés dans l'extraction de l'oxygène de l'eau. Ces bouleversements paraissent donc eux aussi reliés à l'évolution des teneurs en dioxygène : un organisme aérobie épais, dont certaines cellules ne sont plus en contact direct avec l'extérieur, n'aurait pu apparaître tant que l'oxygénation du milieu ne permettait pas d'alimenter efficacement les couches cellulaires internes (figure 18). Ces étapes de diversification pourraient donc être liées au dépassement de valeurs-seuil de l'oxygénation de l'eau, qui aurait « ouvert » de nouvelles voies évolutives, impliquant l'acquisition de tailles plus grandes et d'organisations plus complexes.
Source - © 2009 Payne et al. [27], modifié
Figure 19. Taille des plus grands fossiles au cours de l'histoire de la Terre, depuis 1,9 Ga
Les teneurs en dioxygène sont exprimées en pourcentage de la valeur actuelle (PAL : Présent Atmospheric Level). Triangles rouges : procaryotes ; cercles jaunes : protistes ; carrés bleus ; animaux ; losanges verts : plantes vasculaires ; carrés gris : vendobiontes.
Cette courbe, publiée en 2009 n'intègre pas les « fossiles » gabonais. Nous avons sur-ajouté la tranche d'âge des fossiles gabonais vers -2,1 à –2,2 Ga, ainsi que le volume moyen de ces organismes estimé d'après les dimensions données par l'article de Nature [25]. Les organismes gabonais se trouvent au niveau de la zone de taches bleues, pas très éloignés de Grypania et des cercles jaunes (les protistes).
Les tailles maximales sont illustrées séparément pour les eucaryotes unicellulaires, les animaux et les plantes vasculaires pour l'Ediacarien et le Phanérozoïque. La ligne pleine souligne la tendance de la taille maximale moyenne pour l'ensemble du vivant. L'accroissement de cette valeur s'effectue en deux phases distinctes, approximativement corrélées aux augmentations des teneurs atmosphériques en dioxygène au milieu du Protérozoïque et entre l'Ediacarien et l'Ordovicien ancien.
Références
Fossiles de Burgess et Chengjiang
- [1] J.B. Caron, R.R. Gaines, M.G. Mángano, M. Streng, A.C. Daley, 2010. A new Burgess Shale–type assemblage from the “thin” Stephen Formation of the southern Canadian Rockies. Geology, 38, 9, 811-814. doi:10.1130/G31080.1
- [2] J. Vannier, I. Calandra, C. Gaillard, A Żylińska, 2010. Priapulid worms: Pioneer horizontal burrowers at the Precambrian-Cambrian boundary. Geology, 38, 8, 711-714. doi:10.1130/G30829.1
- [3] P. Van Roy, P. J. Orr, J.P. Botting, L.A. Muir, J. Vinther, B. Lefebvre, K. el Hariri, D.E.G. Briggs, 2010 . Ordovician faunas of Burgess Shale type. Nature 465, n° 7295, 215-218, 2010. doi:10.1038/nature09038
- [4] J. Vannier, 2009. L'Explosion cambrienne ou l'émergence des écosystèmes modernes. C. R. Palevol 8, 133-154. doi:10.1016/j.crpv.2008.10.006
- [5] S.E. Gabbott, J. Zalasiewicz, D. Collins, 2008. Sedimentation of the Phyllopod Bed within the Cambrian Burgess Shale Formation of British Columbia.J. Geol. Soc., 165, 1, 307-318. doi:10.1144/0016-76492007-023
- [6] C.R. Marshall, 2006. Explaining the Cambrian “Explosion” of animals. Annual Review of Earth and Planetary Science 34, 355-384. doi:10.1146/annurev.earth.33.031504. 103001
- [7] K.J. Peterson, M.A. McPeek, D.A.D. Evans, 2005. Tempo and mode of early animal evolution : inferences from rocks, Hox, and molecular clocks. Paleobiology, 31, sp5. 36-55. doi:10. 1666/0094-8373(2005)031[0036:tamoea]2.0.co;2
Communautés fossiles type Ediacara
- [8] S. Xiao S., M. Laflamme,2008. On the eve of animal radiation : phylogeny, ecology and evolution of the Ediacara biota. Trends in Ecology and Evolution, 24, 1, 31-40. doi:10.1016/j.tree.2008.07.015
- [9] G J.H. McCall, 2006. The Vendian (Ediacaran) in the geological record: Enigmas in geology's prelude to the Cambrian explosion. Earth-Science Reviews, 77, 1-229. doi:10.1016/j.earscirev.2005.08.004
- [10] G.M. Narbonne, 2005. The Ediacara biota : neoproterozoic origin of animal and their ecosystems. Annual Review of Earth and Planetary Science, 33, 421-442. doi:10.1146/annurev.earth.33.092203.122519
- [11] M. Brasier, J. Antcliffe, 2004. Decoding the Ediacaran Enigma, Science, 305, 5687, 1115-1117. doi:10.1126/science.1102673
- [12] M.A. Fedonkin, B.M. Waggoner, 1997. The late precambrian fossil Kimberella is a mollusc-like bilaterian organism. Nature, 388, 6645, 868-871. doi:10.1038/42242 doi:10.1038/ 42242
- [13] M.F. Glaessner, 1959. Precambrian Coelenterata from Australia, Africa and England. Nature, 183, 4673, 1472-1473. doi:10.1038/1831472b0
- [14] A. Seilacher, 1992. Vendobionta and Psammocorallia : lost constructions of Precambrian evolution. J. Geol. Soc., 149, n°4, 607-613, 1992. doi:10.1144/gsjgs.149.4.0607
- [15] K.J. Peterson, 2008. The ediacaran emergence of bilaterians : congruence between the genetic and the geological fossil records. Phil. Trans. R. Soc. B: Biological Sciences, 363, 1496, 1435-1443. doi:10.1098/rstb.2007.2233
- [16] S. Xiao, Y. Zhang, A.H. Knoll, 1998. Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite. Nature, 391, 6667, 553-558. doi:10.1038/35318
- [17] L. Yin, M. Zhu, A.H. Knoll, X. Yuan, J. Zhang? J. Hu, 2007. Doushantuo embryos preserved inside diapause egg cysts. Nature, 446, 7136, 661-663. doi:10.1038/nature05682
- [18] J.-Y. Chen, D.J. Bottjer, E.H. Davidson, S.Q. Dornbos, X. Gao, Y.-H Yang, C.-W. Li, G. Li, X.-Q. Wang, D.-C. Xian, H.-J. Wu, Y.-K. Hwu, P. Tafforeau,2006. Phosphatized polar lobe-forming embryos from the Precambrian of southwest China, Science, 312, 5780, 1644-1646. doi:10.1038/10.1126/science.1125964
- [19] D. Condon, M. Zhu, S. Bowring, W. Wang, A. Yang, Y. Jin, 2005. U-Pb ages from the Neoproterozoic Doushantuo Formation, China. Science, 308, 5718, 95-98. doi:10.1126/science.1107765
- [20] P.C.J. Donoghue, S. Bengtson, X.-p. Dong, N.J. Gostling, T. Huldtgren, J.A. Cunningham, C. Yin, Z. Yue, F. Peng, M. Stampanoni, 2006. Synchrotron X-ray tomographic microscopy of fossil embryos. Nature, 442, 7103, 680-683. doi:10.1038/nature04890
- [21] S. Bengston, B. Rasmussen, B. Krapež, 2007. The Paleoproterozoic megascopic Stirling biota. Paleobiology, 33, 3, 351-381. doi:10.1666/04040.1
- [22] B. Rasmussen, S. Bengtson, I.R. Fletcher, N.J. McNaughton, 2002. Discoidal Impressions and trace-like fossils more than 1200 million years old. Science, 296, 5570, 1112-1115. doi:10.1126/science.1070166
- [23] A.Seilacher, P.K. Bose, F. Pflüger, 1998. Triploblastic animals more than 1 billion years ago: Trace fossil evidence from India. Science, 282, 5386, 80-83. doi:10.1126/science.282.5386.80
- [24] T.M. Han, B. Runnegar, 1992. Megascopic eukaryotic algae from the 2.1-billion-yearold Negaunee iron-formation, Michigan. Science, 257, 5067, 232-235. doi:10.1126/10.1126/science.1631544
- [25] A. El Albani, S. Bengtson, D.E. Canfield, A. Bekker, R. Macchiarelli, A. Mazurier, E.U. Hammarlund, P. Boulvais, J.-J. Dupuy, C. Fontaine, F.T. Fürsich, F. Gauthier-Lafaye, P. Janvier, E. Javaux, F. Ossa Ossa, A.-C. Pierson-Wickmann, A. Riboulleau, P. Sardini, D. Vachard, M. Whitehouse, A. Meunier, 2010. Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago. Nature, 466, 7302, 100-104. doi:10.1038/nature09166
- [26] A. Maxmen, 2010. Ancient macrofossils unearthed in West Africa. Nature news, 30 juin 2010. doi:10.1038/news.2010.323
- [27] J.L. Payne, A.G. Boyer, J.H. Brown, S. Finnegan, M. Kowalewski, R.A. Krause Jr, S.K. Lyons, C.R. McClain, D.W. McShea, P.M. Novack-Gottshall, F.A. Smith, J.A. Stempien, S.C. Wang, 2009. Two-phase increase in the maximum size of life over 3.5 billion years reflects biological innovation and environmental opportunity. PNAS, 106, 1, 24-27. doi:10.1073/pnas.0806314106

