Article | 01/10/2006
L'enfant australopithèque de Dikika
01/10/2006
Résumé
Un nouveau fossile d'Australopithèque, et le contexte géologique du site de sa découverte, viennent d'être décrits dans deux articles de la revue Nature du 21 septembre 2006. Ce fossile, daté de 3,31 à 3,35 Ma, ne représente pas une forme nouvelle, puisqu'il est attribué à une espèce déjà bien connue, Australopithecus afarensis (comme la fameuse Lucy), mais il constitue pourtant une découverte exceptionnelle, car il s'agit d'un spécimen juvénile exceptionnellement bien préservé. Il fournit de nombreux indices concernant la locomotion de cette espèce d'australopithèques et son développement ontogénétique. Par ailleurs, la région des Afars, d'où provient ce fossile, reste un lieu de fouille majeur concernant les Australopithèques, et a livré récemment les restes d'une autre espèce, Australopithecus anamensis, connue jusqu'alors au Kenya.
Table des matières
Ce texte a été aimablement relu par Franck Guy (LGBPH de l'Université de Poitiers). Le site "Planet-Vie" publie aussi un article "un coup de jeune chez les afarensis".
Plusieurs découvertes de fossiles attribués au rameau humain (le groupe monophylétique contenant Homo sapiens après séparation de la branche des Chimpanzés) sont venues compléter nos connaissances et relancer les recherches sur l'histoire de notre lignée. Mais il reste exceptionnel de disposer de plusieurs exemplaires d'âges différents d'une même espèce et plus encore d'obtenir des restes du squelette post-crânien de représentant juvénile d'une autre espèce d'Hominine que la nôtre. C'était le cas pour Homo neanderthalensis, l'un de nos cousins les plus récent. Ce l'est désormais aussi pour une autre espèce plus éloigné de nous, l'australopithèque Australopithecus afarensis.
Le nouveau fossile décrit dans la revue Nature du 21 septembre [1][2] et attribué à cette espèce bénéficie, qui plus est, d'un état de conservation exceptionnel. Il donne ainsi accès à plusieurs caractéristiques morphologiques du juvénile d'A. afarensis que l'on peut comparer à leurs homologues chez le juvénile de l'Homme et du Chimpanzé.
Même s'il est loin de pouvoir fournir des réponses définitives, ce fossile, daté de 3,31 à 3,35 Ma, apporte des éléments importants à plusieurs débats actuels sur la locomotion et le mode de vie des Australopithèques et, par conséquent, sur ce qui pouvait distinguer notre propre genre Homo et le genre Australopithecus, deux groupes qui ont coexisté en Afrique pendant plusieurs millions d'années.
Le fossile et son contexte
La découverte
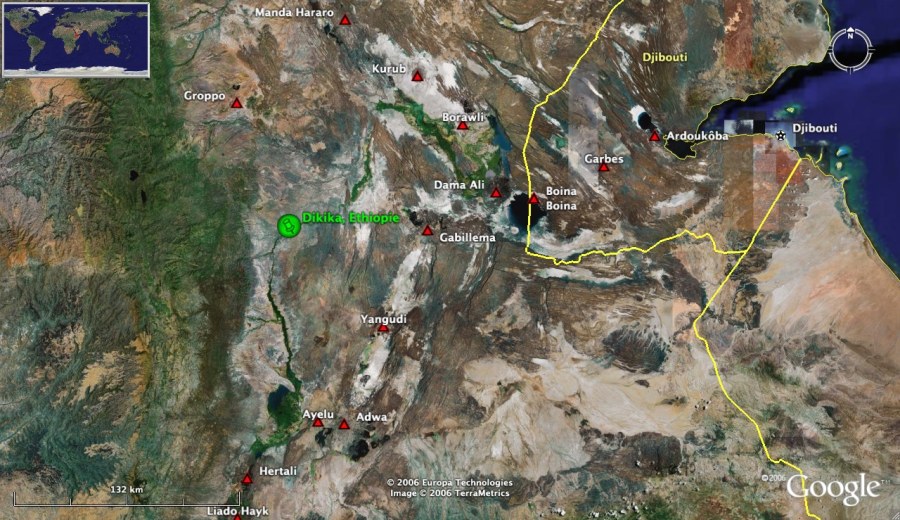
Le fossile nouvellement décrit provient de la localité de Dikika, dans la région des Afars, en Ethiopie [2]. Ce site se situe au sud de la rivière Awash et à proximité de plusieurs autres localités, comme Hadar, où avaient été précédemment exhumés d'autres restes d'hominines, en particulier la grande majorité (90 %) des pièces attribués à l'espèce A. afarensis [4].
Les éléments de ce fossile ont été découvert entre les années 2000 et 2003, mais ils ont nécessités un très long et très minutieux travail de dégagement avant de pouvoir être décrits et publiés.
Source - © 2006 Google Earth
Le contexte géologique de Dikika, Ethiopie
La région de Dikika est une zone aujourd'hui désertique située dans le système du rift éthiopien dans une zone d'extension Nord Ouest - Sud Est, alors que, immédiatement au Nord-Est, commence la zone de rift de la Mer Rouge, de direction d'ouverture Nord Est - Sud Ouest (voir la localisation ci-dessus). Le choix de cette contrée comme zone de fouille vient de ce qu'elle montre un empilement sédimentaire très complet, les formation d'Hadar (3,8 à 2,9 Ma) et de Busidima (2,7 à moins de 0,6 Ma). L'étude de cet empilement a ainsi permis aux auteurs de reconstituer son histoire sédimentaire et les événements tectoniques qui l'ont affectés, événements liés aux jeux des deux systèmes de rift, éthiopien et de la Mer Rouge. Cette grande épaisseur sédimentaire a aussi permis l'étude des changements paléoenvironnementaux de la région, non seulement par l'examen paléontologique des faunes récupérées, mais aussi par la mesure des valeurs isotopiques du carbone 13 (δ13C) de nodules carbonatés formés lors des épisodes de pédogenèse.
La plus grande partie des éléments du fossile ont été exhumés d'un bloc de grès, déposé dans les chenaux d'une plaine deltaïque en bordure d'un lac. Le fossile présente encore plusieurs os en connexion, et ne montre pas de signe d'altération survenue avant enfouissement, signes que l'apport sédimentaire a eu lieu rapidement, probablement lors d'un épisode de grande crue, mais aussi peu de temps après la mort de l'australopithèque. La faune retrouvée en association avec le fossile d'Australopithèque corrobore cette interprétation d'un milieu deltaïque : elle comprend de nombreux gastéropodes d'eau douce, des restes de poissons, d'hippopotamidés, de crocodiles ou encore de tortues aquatiques.
Les valeurs isotopiques δ13C obtenues sur un paléosol situé juste en dessous du grès qui renfermait l'Homininé suggère que les plantes en C3 dominaient très largement la végétation locale, ce qui suggère un milieu arboré relativement humide sans grands contrastes saisonniers (un contexte favorable aux plantes en C4). Là encore, la faune récupérée soutient cette interprétation : les genres identifiés parmi les fossiles de mammifères retrouvés (Equidés, Bovidés, Cervidés, Suidés, Rongeurs, etc.) représentent une faune de milieu arboré sans épisode de sécheresse marqué. Néanmoins, la présence abondante de restes de mammifères brouteurs (antilope, rhinocéros) et d'éléphants indiquent qu'à l'échelle régionale, les prairies ouvertes devaient représenter une part importante des paysages : la région était donc probablement un milieu mosaïque, mais pas, en tout état de cause, une région de forêt dense. Il s'agissait donc d'un milieu plus ouvert que celui associé à d'autres fossiles plus anciens comme Ardipithecus (5,8 et 4,4 Ma), Australopithecus anamensis (4,2 - ~3,9 Ma) ou Kenyanthropus platyops (3,5 Ma).
Les enseignements du jeune Au. afarensis de Dikika
Sur la base de plusieurs caractères du crâne retrouvé et de sa comparaison avec d'autres fossiles de la région de Hadar, les auteurs de la découverte attribuent fermement ce nouveau fossile à l'espèce Australopithecus afarensis. Ce diagnostic est « sans ambiguïté, car la morphologie faciale de cette espèce est évidente même à ce stade juvénile » [1].
Rappelons d'abord quelques éléments sur cette espèce d'australopithèque — l'une des mieux connue — avant de détailler les informations apportées par ce spécimen.
L'espèce Au. afarensis
De très nombreux restes fossiles (plus de 350 éléments décrits aujourd'hui [4]) ont été attribués à l'espèce A. afarensis, depuis sa définition en 1978. Tous ont été découverts dans le rift africain, depuis l'Éthiopie jusqu'à la Tanzanie, la plupart provenant cependant de la région d'Hadar, au Nord de Dikika, sur la rive opposée de l'Awash. Leurs âges s'échelonnent entre 4,1 et 2,9 Ma. Cette espèce est ainsi l'une des mieux connues, d'autant que l'on dispose de plusieurs éléments du squelette post-crânien, en particulier avec le squelette AL-288 — Lucy.
Australopithecus afarensis se signale par un volume crânien relatif (par rapport à la taille du corps) faible, peu différent de celui du Chimpanzé actuel (de 380 à 430 cm3 pour une taille adulte de 1,10 à 1,35 m selon le sexe), une denture en V aux molaires développées, un prognathisme prononcé et un dimorphisme sexuel relativement marqué, au niveau de la taille du corps mais aussi dans la forme des canines. Les membres supérieurs sont relativement longs par rapport aux membres inférieurs. Le pied ne possède pas de voûte plantaire ; le gros orteil est écarté des autres doigts et les tarses sont courts par rapport à la longueur totale du pied. Ces homininés sont considérés comme bipèdes, marcheurs et sans doute capables de grimper efficacement. Mais on ignore lequel de ces deux modes de locomotion, le grimper ou la marche, représentait le mode de déplacement principal de cette espèce.
Ce nouveau fossile — simplement dénommé DIK-1-1 — apporte encore de nombreuses données supplémentaires, puisqu'il comprend plusieurs éléments squelettiques jusqu'alors inconnus pour cette espèce, que plusieurs os sont encore connectés les uns aux autres et qu'il s'agit d'un très jeune individu.
Le jeune DIK-1-1
La face de DIK 1-1 portait encore plusieurs dents identifiées comme des dents de lait et, par tomographie assistée par ordinateur, la présence de plusieurs dents définitives dans la mâchoire a permis d'attester le jeune âge de ce spécimen. Mais ce seule constat ne suffit pas à attribuer un âge chiffré puisque l'on ne connaît pas précisément le développement ontogénétique des australopithèques. Les auteurs proposent cependant pour DIK-1-1 un âge de 3 ans environ, mais en utilisant un modèle tiré de l'étude du développement dentaire chez les grands singes actuels.
L'attribution d'un âge à DIK-1-1 a ensuite permis de considérer certains caractères importants du fossile, comme le volume endôcranien ou la position du trou occipital (foramen magnum), en fonction de cette âge et, là encore, de comparer l'état de ces caractères à ceux d'un chimpanzé actuel de même « âge dentaire ».
La détermination du volume endocrânien du fossile a cependant demandé d'estimer et de corriger les erreurs d'appréciation lié à la déformation des os lors de la fossilisation. Pour cela, les auteurs ont, à nouveau, eu recours à une calibration à partir des données des grands singes actuels. Ils ont établi une corrélation entre le volume endocrânien et plusieurs mesures de l'endocrâne chez des chimpanzés et des gorilles d'âges variables, et ont ensuite appliqué cette équation au cas de DIK-1-1. Le volume de l'endocrâne ainsi obtenu est compris entre 275 et 330 cm3, soit une valeur « peu différente de celle d'un Pan troglodytes (le Chimpanzé commun) du même âge dentaire » [4]. Mais les auteurs rappellent aussi qu'à cet âge juvénile, les états de développement du volume cérébral des grands singes africains et de l'Homme actuel (exprimé en pourcentage du volume endocrânien adulte) sont assez proches.
L'utilisation de la tomographie a également permis d'étudier la forme des canines et de proposer, du fait du dimorphisme sexuel signalée plus haut, un sexe pour ce fossile : DIK-1-1 représenterait une jeune femelle.
Concernant les éléments du squelette post-crânien, les auteurs discutent entre autre du tibia et de l'omoplate. DIK-1-1 présente sur ces os des caractères qui, pour le tibia rappellent ceux de l'Homme, mais, pour l'omoplate, évoquent plutôt ceux du gorille. Les australopithèques présentent ainsi une mosaïque de caractères qui peuvent, individuellement, évoquer ceux de l'un ou l'autre de leurs parents actuels, mais dont la combinaison est propre à ce genre.
Parmi les os retrouvés figure l'os hyoïde, un os très rarement retrouvé chez les Hominidés, car il n'est pas attaché aux autres os du squelette, et par conséquent absent jusqu'ici du registre fossile des Homininés antérieurs à Homo neanderthalensis. Cet os, situé à la base du larynx et qui soutient le cartilage de la pomme d'Adam chez l'Homme actuel, présente chez les Hominidés deux formes : en « bulle » chez les grands singes africains, en « barre » chez l'Orang-outan et l'Homme. Chez DIK-1-1, cet os est similaire à celui du Chimpanzé ou du Gorille, ce qui conforte l'idée que la forme « bulle » représenterait l'état ancestral de cet os chez les Hominidés.

Source - © 2006 Wikipedia
Figure 2. L'os hyoide humain, en vue antérieure
Le cartilage thyroïde (pomme d'Adam) est attaché aux grandes cornes. Tiré de l'ouvrage de Henri Gray, Gray's Anatomy: Descriptive and Surgical , Royaume-Unis, 1858.
Le jeune de Dikika et la bipédie
Le squelette postcrânien de DIK-1-1 confirme un constat déjà réalisé sur les autres fossiles de cette espèce : une « dichotomie fonctionnelle » entre des membres inférieurs plutôt adaptés à un mode de locomotion bipède et une région supérieure du corps rappelant plutôt celle des grands singes adaptés à l'arboricolie.
Cette morphologie fait l'objet de débats quant à son interprétation fonctionnelle : les traits « arboricoles » des Australopithèques impliquent-ils une utilisation réelle et courante de ce mode de locomotion, ou ne sont-ils qu'une rétention de caractères ancestraux chez des animaux principalement bipèdes ?
Sans qu'il permette de trancher réellement ce débat, DIK-1-1 y apporte quelques éléments nouveaux : l'omoplate retrouvée, par exemple, ressemble plus à celle d'un gorille qu'à celle de l'Homme. Or, chez le gorille, les membres supérieurs assurent, dans la locomotion, le soutien du corps et le déplacement (marche sur les phalanges). Il est impossible d'en conclure que DIK-1-1 se déplaçait également en s'aidant des bras, car la relative diversité des architecture de l'omoplate parmi les Hominoïdes reste « encore mal comprise d'un point de vue fonctionnel »[1].
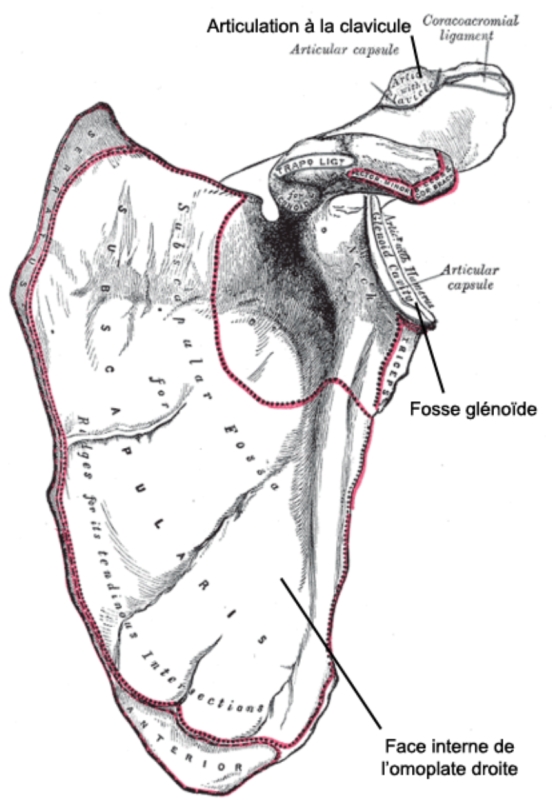
Par contre, la position haute de la fosse glénoïde (cavité de l'omoplate sur laquelle vient s'attacher la tête de l'humérus), constatée sur DIK-1-1, laisse supposer que sa ceinture scapulaire (clavicule et omoplate) était relativement haute par rapport au tronc et à la tête, ce qui est aussi le cas chez les chimpanzés et les gorilles. Chez l'Homme, la position plus basse de cette ceinture permet le découplage des mouvements de la tête et de ceux du tronc. De même, la position de la fosse glénoïde pourrait signifier que DIK-1-1 levait fréquemment les bras au-dessus de sa tête, peut-être pour grimper ou s'accrocher. Une possibilité que suggère aussi le relatif allongement et la courbure des phalanges. La locomotion arboricole pourrait donc bien avoir eu une place relativement importante dans le comportement de cet australopithèque.
Source - © 2006 Wikipedia
Figure 3. Omoplate humaine, face antérieure, montrant la position de la fosse glénoïde
Tiré de l'ouvrage de Henri Gray, Gray's Anatomy: Descriptive and Surgical , Royaume-Unis, 1858.
Conclusions
DIK-1-1, un fossile majeur pour connaître les Au. afarensis
DIK-1-1 ne constitue donc pas, comme d'autres fossiles récemment décrits, un bouleversement des connaissances. Par contre, il vient compléter le registre de l'une des espèces de ce groupe entièrement éteint que constituèrent les australopithèques. Il comprend des éléments squelettiques jusqu'alors absents des autres représentants fossiles de ce genre et permet donc d'affiner les reconstitutions anatomiques de cette espèce. Ce faisant, il vient alimenter les recherches et les débats la concernant, comme par exemple leur mode de locomotion (plus grimpeur arboricole que marcheur ?), mais aussi leur développement ontogénétique ou leur habitat (un milieu arborée dans une région mosaïque de plaines ouvertes et de zones boisées).
Se pose cependant toujours, dans l'interprétation de ces fossiles, le problème de la réelle signification fonctionnelle des caractéristiques anatomiques constatées : on estime encore mal si les différences de forme de certains os (l'omoplate par exemple) entre l'Homme, les australopithèques et les grands singes, par exemple, indiquent des capacités fonctionnelles différentes. Résultent-elles d'adaptations différentes ou simplement d'une diversité morphologique "fonctionnellement neutre" ?
Australopithecus afarensis et Au. anamensis
Quelques mois avant la description de DIK-1-1 a été publié celle d'autres restes, attribués à l'espèce Australopithecus anamensis [5]. Ces fossiles sont eux aussi importants parce qu'ils proviennent de la même zone d'Ethiopie que DIK-1-1 et les autres Australopithecus afarensis : la région de l'Awash moyenne (Middle Awash). Or, jusqu'à présent, les principaux spécimens d'A. anamensis provenaient du Kenya, 1000 km au Sud-est. Qui plus est, c'est aussi d'Ethiopie que provient un fossile encore plus ancien, Ardipithecus ramidus
Par ses caractéristiques anatomiques et son âge (~4,12 Ma), ce fossile est bien rattaché aux A. anamensis, la plus ancienne espèce du genre Australopithecus. De plus, son âge est similaire à celui des plus anciens A. anamensis connus au Kenya. Selon les auteurs [5], l'abondance des restes suggère que cette espèce occupait régulièrement cette région et que ces fossiles ne représentent pas que quelques individus égarés. Ce qui pourrait signifier que l'aire d'origine de cette espèce serait plus éthiopienne que kenyane.
De plus, pour l'instant, on ne connaît pas de fossiles d'A. anamensis et de A. afarensis contemporains dans le temps (les A. anamensis sont datés de 4,2 à 3,9 Ma, les A. afarensis apparaissent à partir de 3,6 Ma), et de même, les espèces Ardipithecus ramidus (4,4 Ma) et Australopithecus anamensis paraissent temporellement, mutuellement exclusives. Retrouver ainsi ces trois espèces dans la même région, et les uns après les autres, vient alimenter le débat sur leurs relations de parenté. Si l'aire de répartition de A. anamensis s'est étendue du Kenya à l'Ethiopie, où se trouvaient auparavant Ardipithecus et où apparaissent les A. afarensis, cela pourrait impliquer :
- une relation de parenté directe entre ces trois espèces, donc une évolution (par cladogenèse ou anagenèse) des A. anamensis à partir des Ardipithecus et de même des A. afarensis à partir des A. anamensis ou d'une sous-population est-africaine de ces derniers.
- ou/et un remplacement des Ardipithèques par les Australopithèques dans cette région.
Les questions posées par ce groupe d'Homininés particulier sont donc tout aussi nombreuses que celles qui restent en suspens à propos de notre propre genre Homo et, dans un cas comme dans l'autre, c'est l'augmentation du registre fossile qui reste seul à même d'apporter des réponses.
Références
- [1] Z. Alemseged, F. Spoor, W.H. Kimbel, R. Bobe, D. Geraads, D. Reed, J.G. Wynn, 2006. A juvenile early hominin skeleton from Dikika, Ethiopia. Nature, 443, 7109, 296-301
- [2] J.G. Wynn, Z. Alemseged, R. Bobe, D. Geraads, D. Reed, D.C. Roman, 2006. Geological and palaeontological context of a Pliocene juvenile hominin at Dikika, Ethiopia. Nature, 443, 7109, 332-336
- [3] Y. Coppens, P. Picq, 2001. Aux origines de l'humanité 1 - De l'apparition de la vie à l'homme moderne. Fayard . ISBN 2-213-60369-3
- [4] É. Crubézy, J. Braga, G. Larrouy, 2002. Anthropobiologie. Masson. ISBN 2-294-00656-9
- [5] T.D. White et al., 2006. Assa Issie, Aramis and the origin of Australopithecus. Nature, 440, 7086, ,883-889